КЛАДОЭНДЕЗИС НАСЕКОМЫХ

![]()
|
КЛАДОЭНДЕЗИС НАСЕКОМЫХ |
|||||||
|
|
|
fg | |||||
IV-1.3.2.2. Chelicerata Heymons 1901 – хелицеровые
(рис. 4.5D)
|
Названия, однозначно соответствующие по циркумскрипции: — Chelicerata Heymons 1901; — Euarachnida Lankester 1904; — Panarachnida Lameere 1933; — Euchelicerata Ax 1984. Название, неоднозначно соответствующее по циркумскрипции (соответствующее также Chelicerophora): — Cephalogradi Crampton 1928b, s.str. По циркумскрипции также соответствует: — «Acerata» sensu Kingsley 1894 (non Lankester 1888). Иерархическое название: Araneus/fg9 [fg:(1757)1758] (sine Pycnogonum; incl. Limulus). |
|
Ранг. Обычно от класса до типа.
Название. Некоторые авторы используют название «Chelicerata» вместо Chelicerophora, т.е. для таксона, включающего Pantopoda. Исходно (Heymons 1901) название Chelicerata было опубликовано в работе, посвященной не классификации членистоногих, а онтогенезу и анатомии сколопендры. Классификация членистоногих в этой работе дана очень кратко; в составе Chelicerata указаны только Xiphosura (с представителем Limulus) и Arachnoidea (с представителем Scorpio). В то время морских пауков (Pantopoda) иногда помещали в состав класса Arachnoidea (или Arachnida), иногда рассматривали отдельно от него. Так что перечень подчиненных таксонов в данном случае не позволяет понять, какая из двух последующих интерпретаций названия Chelicerata соответствует исходной. Однако приведенный в этой работе диагноз таксона Chelicerata дает однозначный ответ на этот вопрос. Согласно этому диагнозу «cephalon» Chelicerata включает 7 метамеров, причем конечности первого метамера утрачены, конечности второго метамера преобразованы в хелицеры, а конечности следующих пяти метамеров являются частью гнатоподами, частью обычными ногами. Исходя из того, что строение Pantopoda было хорошо известно и явно противоречит этому диагнозу, следует считать, что исходно Pantopoda не были включены в состав Chelicerata.
Аутапоморфии Chelicerata
(1) Редукция конечностей задней части тела. Конечности VII посторального сегмента утратили локомоторную функцию [как и следующие за ними – см. Chelicerophora (3)] (тогда как у Pantopoda задняя пара ходильных конечностей относится к сегменту, являющемуся не менее чем VII посторальным). Туловищные сегменты дифференцированы следующим образом (рис. 4.5D).
Все тело подразделено на головогрудь (cephalothorax), или просому (prosoma) – передний отдел, включающий I–VI посторальные сегменты и несущий ротовые и локомоторные конечности, и брюшко (abdomen), или опистосому (opisthosoma) – задний отдел, включающий последующие посторальные сегменты и не несущий локомоторных конечностей. Таким образом, поскольку конечности I посторальной пары преобразованы в хелицеры [см. Chelicerophora (2)], имеется не более 5 пар локомоторных конечностей, расположенных на просоме.
У большинства хелицеровых I–VI сегменты слиты, так что вся дорсальная сторона головогруди представляет собой цельный головогрудной щит – пельтидий (peltidium), не разделенный на тергиты; лишь у Mycetophora, Palpigradi, Tartarides и некоторых Monomerosomata имеется пропельтидий (propeltidium) – общий дорсальный щит I–IV сегментов, тогда как V и VI сегменты головогруди (несущие две последние пары ног) обособлены и могут иметь отдельные тергиты, называемые мезопельтидий (mesopeltidium) и метапельтидий (metapeltidium). В прошлом предполагали, что пропельтидий гомологичен голове трилобитов, которую ошибочно считали четырехсегментной (см. Trilobitomorpha: «Классификация»). В строении пяти пар конечностей II–VI сегментов может проявляться эволюционная корреляция; они могут быть сходны по строению или различны (например, у большинства Arachnida конечности II сегмента, называемые педипальпами, сильно отличаются от других).
VII посторальный сегмент считается промежуточным или I сегментом опистосомы. Его конечности преобразованы в аналог нижней губы – пару хиляриев (chilaria); среди современных хелицеровых хилярии сохраняются только у Xiphosura (рис. 4.5D), у всех Arachnida утрачены.
На опистосоме бывают развиты конечности, не имеющие локомоторной функции – половые крышки на II сегменте опистосомы [т.е. на VIII посторальном сегменте – см. (3)], а также жабры и другие нелокомоторные конечности. Они бывают развиты до VII сегмента опистосомы (т.е. до XIII посторального сегмента) включительно – такое максимальное число конечностей опистосомы имеется у мечехвостов (Xiphosura).
Сегменты начиная с XIV посторального всегда лишены конечностей.
(2) Утрата одной пары глазков. Имеется не более одной пары глазков (в отличие от Pantopoda, у которых сохраняются две пары глазков) [см. Euarthropoda (4)]. Лишь у Xiphosura помимо пары нормально развитых глазков имеется пара светочувствительных органов на вентральной стороне просомы, возможно представляющая собой вестигии второй пары глазков (Paulus 1979). Помимо пары глазков могут сохраняться латеральные глаза, которые бывают фасеточными или состоят из нескольких простых глаз; в последнем случае глазки и глаза могут быть внешне сходными, различаясь только по иннервации (табл. IV.3).
Признаки Chelicerata неясного филогенетического значения
(3) Положение полового отверстия. Половое отверстие всегда находится на вентральной стороне VIII посторального сегмента (тогда как у Pantopoda половые отверстия на туловище утрачены и имеются вторичные парные половые отверстия на ногах). В остальном строение половой системы разнообразно: у Xiphosura гонады, их протоки и половые отверстия парные; у Arachnida гонады самца и/или самки бывают парными или непарными, их протоки парные, а половое отверстие непарное.
(4) Супраанальный придаток. На конце тела дорсальнее анального отверстия имеется подвижно причлененный непарный супраанальный придаток (рис. 4.5D). У разных хелицеровых число сегментов опистосомы различно, и их супраанальный придаток находится на разных по счету сегментах; на основании этого некоторые авторы считают супраанальный придаток либо результатом срастания нескольких последних сегментов опистосомы, либо тельсоном. В отличие от сегментов или тельсона [см. Metameria (1a)] супраанальный придаток не содержит кишки и анального отверстия. Форма супраанального придатка бывает различной: в виде шипа, лопасти, крюка или членистого хвоста; иногда он утрачен.
(5) Эпиморфоз. Развитие эпиморфное: из яйца выходит животное с полным набором сегментов (в отличие от анаморфного развития Pantopoda). У большинства Chelicerata число пар конечностей также не меняется с возрастом; лишь у Xiphosura личинка первого возраста не имеет трех задних пар жаберных конечностей на XI–XIII сегментах, а у Monomerosomata личинка первого возраста не имеет задней пары ног (расположенных на VI сегменте).
Плезиоморфии и изменчивые признаки Chelicerata
(6) Выделительные железы. Парные выделительные железы [см. Gnathopoda (1)] могут сохраняться в просоме [см. (1)]. У Xiphosura (обитающих в морской воде) имеется пара функционирующих желез, каждая из которых включает целомические мешки четырех II–V посторальных сегментов и один общий выводной проток, открывающийся на V посторальном сегменте позади оснований предпоследней пары ног. У различных Arachnida (являющихся наземными и обладающих выделительными мальпигиевыми сосудами – см. «Классификации» III) в одном или нескольких из этих сегментов также бывают развиты парные выделительные железы, открывающиеся одним или двумя парными протоками на II или III и/или V посторальном сегменте (Snodgrass 1938).
Геологический возраст. Ордовик – ныне (рис. 2.2); среди известных представителей наиболее древними являются Gigantostraca (см. ниже).
Состав и распространение. Более 100 000 видов (Zhang 2011); всесветно.
Классификации Chelicerata. Все предложенные к настоящему времени классификации и гипотезы о филогении хелицеровых внутренне противоречивы. Поэтому здесь отдельно даны характеристики наиболее признанных таксонов (см. I), отдельно – обсуждение филогенетических связей некоторых из них (см. II–VIII).
I. Деление Chelicerata на Xiphosura, Gigantostraca, Toxicura, Pseudoscorpii, Mycetophora, Phalangium/fg1, Palpigradi, Caulogastra, Ricinulei и Monomerosomata.
1. Xiphosura Latreille 1802 – мечехвосты (рис. 4.5D) {synn.circ.: Xiphura Leach 1814 = Acera Blainville 1816 = Merostomata Dana 1846 = Polychelata Börner 1902; nom.typ: Limulus/fg [f:1820; g:1785]}.
Голофилетический таксон, характеризующийся следующими аутапоморфиями. Вся просома, или головогрудь [слитная до VI посторального сегмента включительно – см. Chelicerata (1)] дорсально образует широкий выпуклый щит, закругленный спереди и с заостренными заднебоковыми углами; глаза и глазки находятся на дорсальной стороне этого щита. VIII–XIV посторальные сегменты (образующие опистосому) неподвижно слиты друг с другом, а VII посторальный сегмент (несущий хилярии) образует подвижное сочленение просомы с опистосомой. Таким образом, общее число сегментов тела значительно меньше, чем у многих других Chelicerata, имеющих до 17–19 посторальных сегментов. Супраанальный придаток [см. Chelicerata (4)] представляет собой длинный шип, подвижно сочлененный с опистосомой.
При этом мечехвосты представляют собой наиболее архаичную группу хелицеровых. Сохраняется пара фасеточных глаз и пара простых глазков [единственная – см. Chelicerata (2)]. Имеется наиболее полный среди Chelicerata набор конечностей на всех I–XIII посторальных сегментах. Расположенные на просоме конечности II–VI посторальных пар [см. Chelicerata (1)] сходны по строению (не дифференцированы на педипальпы и ноги) и причленены вентрально близко друг к другу, окружая ротовое отверстие; каждая конечность сохраняет медиальный жевательный выступ первого членика (гнатобазу) и развитый ходильный телоподит, т.е. представляет собой примитивный гнатопод [см. Euarthropoda (3)]; у конечности VI пары первый членик, кроме того, несет латеральный придаток, возможно гомологичный наружной ветви трилобитов [ср.: Trilobitomorpha (1)]. Каждая ходильная конечность заканчивается клешней, наружный подвижный палец которой образован простым когтевидным претарсусом [имеющим двухмыщелковое сочленение – см. Chelicerophora (4)], а внутренний неподвижный палец – вершинным отростком тарсуса; от основания претарсуса (подвижного пальца) отходят антагонистические мышцы – флексор и экстензор, крепящиеся внутри тарсуса [то же у Pantopoda, в отличие от всех прочих современных Chelicerata – см. ниже, III(3)]. На VII посторальном сегменте имеется пара одночлениковых хиляриев; они расположены между основаниями ног VI сегмента, т.е. непосредственно позади гнатобаз, и составляют часть ротового аппарата (у всех прочих Chelicerata эти конечности полностью утрачены). На VIII посторальном сегменте, т.е. на первом сегменте слитной части опистосомы, имеется пара половых крышек; на IX–XIII посторальных сегментах имеется 5 пар жаберных придатков, служащих для дыхания в морской воде. Среди современных Chelicerata только мечехвосты сохраняют первичный морской образ жизни. Крупные (значительно крупнее наземных хелицеровых), длина тела 500–900 мм.
В современной фауне имеется лишь 5 видов Xiphosura: один вид (L. polyphemus) распространен у атлантического побережья Северной Америки, 4 вида в морях Юго-Восточной Азии. В ископаемом состоянии достоверно известны начиная с нижнего силура (Mickulic & al. 1985).
2. †Gigantostraca Haeckel 1866 – ракоскорпионы {synn.circ.: Pomatostoma Thorell & Lindstrom 1885 = Haplochelata Börner 1902; nom.typ.: Eurypterus/fg [f:1845; g:1825]}. Вымершие палеозойские животные, существовшие с ордовика до перми; судя по всему, сохраняли первичный морской образ жизни. Конечности просомы по-разному специализированы у разных представителей: конечности всех II–VI пар бывают ходильными, иногда конечности II–III пар хватательные, иногда конечности VI пары расширенные, плавательные. Опистосома заканчивается XIX сегментом и бывает подразделена на более широкую 7-сегментную мезосому и более узкую 5-сегментную метасому, на конце которой находится супраанальный придаток (то же у Toxicura); форма супраанального придатка бывает разнообразной, от шиповидной (как у Xiphosura) и когтевидной (как у Toxicura) до лопатковидной. Имеются фасеточные глаза и пара медиальных глазков (то же у Xiphosura и некоторых вымерших Toxicura). Длина тела 100–1800 мм.
Возможно, Gigantostraca – парафилетический таксон, предковый для Toxicura или в целом для Arachnida (см. II и III).
3. Toxicura Cook 1899b – скорпионы {synn.circ.: Ctenophora Pocock 1893c (non Ctenophorae Eschscholtz 1829) = Cteidophora Börner 1902 = Pectinifera Lankester 1910; nom.hier: Scorpio/fg1 [f:1802; g:L.1758(gen.238)]}. Просома дорсально образует цельный щит. Хелицеры сохраняют 3-члениковое клешневидное строение. Конечности II посторальной пары (педипальпы) утратили претарсус и преобразованы в клешни, не гомологичные клешням Xiphosura (то же у Pseudoscorpii); их коксы утратили гнатобазы (в отличие от большинства Chelicerata). Конечности III–VI пар служат ногами; ноги всех четырех пар направлены вершинами вперед, коленями назад; коксы двух передних пар ног сомкнутые, подвижные и несут гнатобазы (жевательные выступы, направленные вперед); коксы двух задних пар ног также несут направленные вперед выступы, но неподвижные и образуют значительную часть вентральной стенки тела (так что стернит развит лишь в задне-медиальной части просомы). Промежуточный сегмент (VII посторальный) редуцирован. Опистосома [см. Chelicerata (1)] заканчивается XIX посторальным сегментом (т.е. является наиболее длинной среди современных Chelicerophora); первые 7 сегментов опистосомы (посторальные сегменты VIII–XIV), называемые мезосомой (mesosoma), не утоньшены; последние 5 сегментов опистосомы (посторальные сегменты XV–XIX) образуют тонкую гибкую метасому (metasoma) (то же у вымерших Gigantostraca). Супраанальный придаток [см. Chelicerata (4)] представляет собой крюк с ядовитой железой. Сохраняются конечности VIII и IX посторальных сегментов: на VIII сегменте пара половых крышек (то же у Xiphosura); на IX сегменте пара гребневидных органов (уникальное образование). У современных наземных скорпионов на вентральной стороне мезосомы имеются 4 пары книговидных легких, расположенных по паре на X–XIII посторальных сегментах [см. III: Arachnida (5b)]; у вымерших силурийских и девонских морских скорпионов легочных отверстий нет. Длина тела 50–200 мм.
Все современные скорпионы обитают на суше. Современных и вымерших наземных скорпионов объединяют в таксон Dionychopodes Thorell & Lindström 1885 {nom.hier.: Scorpio/fg2 [f:1802; g:1758]}, который характеризуется наличием двух коготков на претарсусе каждой ходильной ноги [см. III: Arachnida (3)]. У всех современных скорпионов глаза не фасеточные: имеется до 5 пар однолинзовых глаз по бокам просомы и пара глазков на медиальном бугорке (у некоторых представителей число глаз уменьшено, у некоторых глаза и глазки утрачены). Dionychopodes известны начиная с девона; в современной фауне более 2000 видов (Zhang 2011); во всех частях света, преимущественно в тропиках и субтропиках.
Кроме того, известны вымершие силурийские морские скорпионы; объединяют в таксон Apoxipodes Thorell & Lindström 1885 {nom.typ: Palaeophonus/fg [fg:1885]}, который характеризуется заостренными претарсусами без двух боковых коготков. Некоторые из этих морских скорпионов имеют вполне развитые латеральные фасеточные глаза и пару глазков.
4. Pseudoscorpii Siebold 1848 – ложноскорпионы {synn.circ.: Pseudoscorpioda Haeckel 1866 = Pseudoscorpionea Gegenbaur 1878 = Pseudoscorpionina Lankester 1881 = Diplochelonethi Thorell 1882 = Pseudoscorpionida Lankester 1888 = Pseudoscorpionidea Lang 1888 = Chelata Petrunkevitch 1946 = Pseudoscorpionoidea Beklemishev 1952 = Pseudoscorpionodea Dubinin 1962 (non Pseudoscorpiones Latreille 1825); nom.typ.: Chelifer/fg [f:1826; g:1762]}. Названия Pseudoscorpii и др. не следует путать с названием Pseudoscorpiones, которое является старшим синонимом Haplocnemata (см. VII).
Просома дорсально образует цельный щит. Глазки утрачены; имеется до 2 пар однолинзовых глаз по бокам просомы (у многих представителей задняя пара утрачена, иногда утрачены обе пары). Каждая хелицера [исходно трехчлениковая клешня – см. Chelicerophora (2)] не имеет проксимального членика, с вентральным подвижным пальцем (то же у Mycetophora – см. ниже); на вершине подвижного пальца открывается проток паутинной железы, расположенной в просоме. Конечности II пары (педипальпы) утратили претарсус и преобразованы в клешни, не гомологичные клешням Xiphosura (то же у Toxicura – см. VI); их коксы с гнатобазами. Конечности III–VI пар служат ногами; ноги двух передних пар направлены вперед, ноги двух задних пар направлены назад; коксы всех ног без гнатобаз, сомкнуты (так что стерниты просомы не развиты); претарсус с двумя коготками. Опистосома заканчивается XVIII посторальным сегментом (т.е. состоит из 12 сегментов, включая прегенитальный), не подразделена на мезосому и метасому, без каких-либо конечностей и без супраанального придатка. Все представители наземные; имеют трахейную систему, открывающуюся двумя парами дыхалец на IX и X посторальных сегментах. Мелкие, длина тела 2–7 мм. Известны начиная с девона; в современной фауне около 3500 видов (Zhang 2011); во всех частях света, преимущественно в тропиках и субтропиках.
5. Mycetophora Pocock 1893c – сольпуги, или фаланги {synn.circ.: Malleolifera Petrunkevitch 1946 = Solifugomorpha Dubinin 1956); nom.typ.: Solpuga/f=Galeodes/g [f:1815; g:1791]}. Аутапоморфия: ноги последней (VI) пары со стебельчатыми треугольными или Т-образными придатками (malleoli) – на каждой ноге обычно имеется пять таких придатков, по два на первых двух члениках и один на третьем членике. Просома дорсально подразделена на пропельтидий и подвижные тергиты V и VI сегментов [см. Chelicerata (1)]. Глаза утрачены, имеется пара глазков (то же у Opiliones). Хелицеры сильно увеличены; каждая хелицера [исходно трехчлениковая клешня – см. Chelicerophora (2)] не имеет проксимального членика, с вентральным подвижным пальцем (то же у Pseudoscorpii). Конечности II пары (педипальпы) крупные, похожи на ноги, но с втяжным пузырьком вместо претарсуса; к пузырьку подходят две мышцы, как и к претарсусу ноги. Коксы педипальп с гнатобазами (то же у Pseudoscorpii, Phalangium/fg1, Caulogastra, Monomerosomata). У конечностей III–VI пар претарсус с двумя коготками; конечности III пары не используются при ходьбе, а служат только для осязания (то же у Pedipalpi и Palpigradi); конечности IV–VI пар служат ногами; коксы всех конечностей III–VI пар без гнатобаз (то же у Pseudoscorpii, Caulogastra, Monomerosomata), но крупные и сомкнутые, так что стерниты просомы не развиты. Опистосома заканчивается XVII сегментом (т.е. состоит из 11 сегментов, включая прегенитальный), без конечностей и без супраанального придатка. Все представители наземные; имеют трахейную систему, открывающуюся дыхальцами на IX–XI посторальных сегментах. Длина тела 10–70 мм.
Известны начиная с карбона; в современной фауне более 1000 видов (Zhang 2011); во всех частях света, преимущественно в тропиках и субтропиках.
6. Phalangium/fg1 [f:1802; g:L.1758(gen.236)] (incl. Siro) – сенокосцы. Аутапоморфии: (1) по бокам просомы имеется пара отверстий пахучих желез; (2) на генитальном (VIII посторальном) сегменте имеется выдвигающийся вперед трубчатый вырост с половым отверстием на вершине; у самки этот вырост служит яйцекладом, а у самца – пенисом и используется для внутреннего оплодотворения (в отличие от сперматофорного оплодотворения у других хелицеровых). Просома дорсально образует цельный щит. Хелицеры сохраняют 3-члениковое клешневидное строение. Конечности II пары (педипальпы) щупикообразные или хватательные (у Opiliones-Laniatores); оканчиваются таким же коготком, как ноги следующих пар. Конечности III–VI пар ходильные; претарсус в виде одного коготка (лишь у Laniatores на ногах двух последних пар по два коготка). Опистосома заканчивается XVII сегментом, без конечностей и без супраанального придатка. Все представители наземные; имеют трахейную систему, открывающуюся парой дыхалец на вентральной стороне VIII посторального сегмента по бокам от половой крышки. Длина тела 1–20 мм. Известны начиная с карбона. Таксон Phalangium/fg1 делится на Anepignathi и Opiliones.
6.1. Anepignathi Thorell 1882 {nom.typ.: Siro/fg [f:1839; g:1796] (incl. Stylocellus)}; включает Cyphophthalmus/fg [fg:1868]. Отверстия пахучих желез по бокам просомы открываются на паре дорсальных выступов. Глазки утрачены, бывает развита пара глаз по бокам просомы; многие виды слепые. Около 200 видов (Zhang 2011).
6.2. Opiliones Sundevall 1833 {nom.hier.: Phalangium/fg2 [f:1802; g:1758]}. Анальное отверстие смещено вентрально. Генитальный вырост прикрыт крышкой, образованной следующим (XI) стернитом. Ноги бывают сильно удлинены. Глаза утрачены, имеется пара глазков на медиальном выступе (то же у Mycetophora). Около 6500 видов (Zhang 2011); во всех частях света. Opiliones делят на Laniatores Thorell 1876 {nom.typ.: Gonyleptes/fg [f:1833; g:1818]} и Palpatores {nom.hier.: Phalangium/fg3 [f:1802; g:1758]}.
7. Palpigradi Thorell 1888 {synn.circ.: Microthelyphonida Grassi 1885 = Latisterna Petrunkevitch 1949; nom.typ: Eukoenenia/fg [f:1901; g:1901]}. Просома дорсально подразделена на пропельтидий и отдельные тергиты V и VI сегментов [см. Chelicerata (1)]. Глазки и глаза утрачены. Хелицеры сохраняют 3-члениковое клешневидное строение. Конечности II–VI пар сходного строения, все без гнатобаз (в отличие от прочих Chelicerata), у всех претарсус с тремя коготками; конечности II пары (педипальпы) и IV–VI пар служат ногами, а конечности III пары – осязательными органами. Имеется суженный VII (промежуточный) сегмент. Опистосома заканчивается XVII сегментом, несет многочлениковый супраанальный придаток [см. Chelicerata (4)] (то же у Caulogastra-Uropygi). На VIII–XII посторальных сегментах по паре втяжных пузырьков, вероятно, гомологичных конечностям. Все представители наземные. Мелкие, длина тела 0,5–3 мм. В современной фауне более 80 видов (Zhang 2011); в тропиках и субтропиках всех частей света; в ископаемом состоянии известны из юры.
8. Caulogastra Pocock 1893c {synn.circ.: Arachnidea Hammen 1977 = Tetrapulmonata Shultz 1990; nom.typ.: Araneus/fg [fg:(1757)1758] (incl. Phrynus)}. Аутапоморфии: (1) каждая хелицера [исходно трехчлениковая клешня – см. Chelicerophora (2)] представляет собой 2-члениковую полуклешню: первый членик утрачен, а неподвижный палец второго членика укорочен или утрачен; (2) аксонема сперматозоида имеет 9+3 микротрубочек [вместо обычных 9+2 – см. Eucaryota (2b)]. Имеется до 3 пар однолинзовых глаз и пара глазков (у Amblypygi и Oxopoei глаза образуют пару компактных групп по 3 глаза в каждой – т.е. сохраняют строение, сходное с фасеточным; у Maripalpi глаза разрозненны; у Tartarides глаза и глазки утрачены). Коксы конечностей II пары (педипальпы) либо с гнатобазами (у Maripalpi, Amblypygi и Tartarides), либо слиты медиально (у Oxopoei); остальные коксы без гнатобаз. У конечностей III–VI или IV–VI пар (ходильных ног) претарсус с 3 или 2 коготками. Опистосома заканчивается XVIII сегментом; супраанальный придаток имеется у Uropygi, отсутствует у Maripalpi и Amblypygi (см. ниже). Все представители наземные. Исходно на VIII и IX посторальных сегментах имеется по паре легких [см. III: Arachnida (5b)]; иногда легкие задней пары или обеих пар утрачены или заменены трахеями. Caulogastra делятся на Maripalpi и Pedipalpi.
8.1. Maripalpi Cook 1899b – пауки {syn.circ.: Diechisoma Billberg 1820; nom.typ: Araneus/fg [fg:(1757)1758] (sine Phrynus; incl. Liphistius)}. Подвижный палец хелицеры [двучлениковой – см. Caulogastra] у вершины с отверстием ядовитой железы, которая расположена либо в пределах хелицеры (в примитивном случае), либо вдается в полость просомы. У половозрелых самцов конечности II пары (педипальпы) модифицированы и используются для копуляции. Конечности X–XI посторальных сегментов преобразованы в две пары прядильных придатков, несущие отверстия паутинных желез и обычно называемых «паутинные бородавки». Длина тела 0,8–110 мм. Около 44 000 видов (Zhang 2011); всесветно.
8.2. Pedipalpi Latreille 1806 {syn.circ.: Pedipalpia Rafinesque 1815; nom.hier.: Thelyphonus/f1=Phrynus/g1 [f:1835; g:1801]; старшее название спорного статуса: Tarantula/fg [f:1814; g:1793]}. Конечности II пары (педипальпы) хватательные. Конечности III пары (соответствующие передним ногам других паукообразных) сильно удлинены, жгутовидные, без коготков, выполняют функцию антенн или щупиков. Pedipalpi делятся на Amblypygi и Uropygi. Некоторые авторы не признают таксон Pedipalpi и объединяют Maripalpi и Amblypygi в таксон Labellata Petrunkevitch 1949.
8.2.1. Amblypygi Thorell 1882 {nom.hier.: Phrynus/f1=g2 [f:1844; g:1801]}. Супраанальный придаток отсутствует. Известны начиная с карбона; в современной фауне около 200 тропических видов.
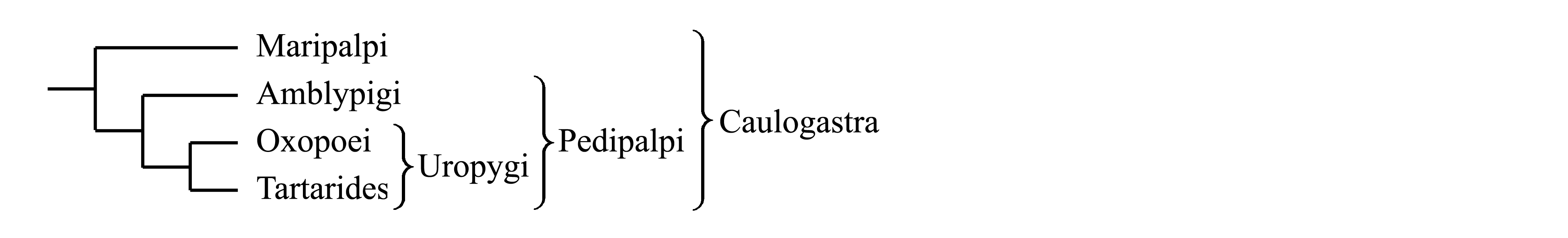
8.2.2. Uropygi Thorell 1882 {nom.hier.: Thelyphonus/f2=g1 [f:1835; g:1802] (incl. Hubbardia)}. Имеется нитевидный многочлениковый или нерасчлененный супраанальный придаток [см. Chelicerata (4)]; то же у Palpigradi. Uropygi делятся на Oxopoei и Tartarides.
8.2.2.1. Oxopoei Thorell 1888 {synn.circ.: Oxypoei auct. = Holopeltidia Börner 1904b; nom.hier.: Thelyphonus/f3=g2 [f:1835; g:1802]}. Длина тела до 750 мм. Известны начиная с карбона; в современной фауне более 100 видов (Zhang 2011); в Ориентальной и Неотропической областях.
8.2.2.2. Tartarides Cambridge 1872 {synn.circ.: Colopyga Cook 1899b = Schizopeltidia Börner 1904b; nom.typ.: Hubbardia/fg [fg:1899]}. Просома дорсально подразделена на пропельтидий и подвижные тергиты V и VI сегментов [тогда как у всех остальных Caulogastra просома дорсально образует цельный щит — см. Chelicerata (1)]. Длина тела 2–7 мм. более 250 тропических видов (Zhang 2011); в ископаемом состоянии известны из палеогена.
9. Ricinulei Thorell 1876b {synn.circ.: Rhignogastra Cook 1899b = Cucullata Pocock 1929; nom.typ.: Ricinoides/f=Cryptocellus/g [f:1929; g:1874]}. Личинка первого возраста лишь с 3 парами ног (то же у Monomerosomata). Более 70 видов (Zhang 2011).
10. Monomerosomata Leach 1815b – клещи {synn.circ.: Holetria Rafinesque 1815 = Phthiridia Rafinesque 1815 = Sympagisoma Billberg 1820 = Hologastres Duges 1834 = Sphaeracara Haeckel 1889 = Rhynchostomi Lankester 1910; nom.hier.: Acarus/fg1 [f:1802, g:L.1758(gen.235)]}. Имеется подвижная гнатосома (gnathosoma), образованная слитыми коксами педипальп (конечностей II пары); во многих случаях гнатосома крупная и охватывает основания хелицер. Личинка первого возраста лишь с 3 парами ног: в отличие от имаго она не имеет конечностей VI посторальной пары; у некоторых клещей в онтогенезе поступательно добавляется последняя пара ног и задние сегменты опистосомы – то есть развитие напоминает примитивный анаморфоз; у Peritremata в эмбриогенезе развиваются все четыре пары ног, но затем последняя пара уменьшается и втягивается в туловище, и из яйца выходит шестиногая личинка, которая при последующей линьке снова становится восьминогой – то есть развитие принципиально отличается от анаморфоза. Строение клещей разнообразно. Просома дорсально либо расчленена, либо образует цельный щит, либо слита с опистосомой; у многих клещей все туловище нерасчлененное (с чем связано название «Monomerosomata»). Хелицеры могут сохранять 3-члениковое клешневидное строение, либо модифицированы. Конечности II пары [педипальпы – см. III: Arachnida (2)] обычно щупикообразные. Конечности III–VI пар (ноги) без гнатобаз; претарсус с одним или двумя коготками. Сегменты опистосомы обособлены или слиты, без супраанального придатка, без конечностей или с генитальными придатками на VIII–X посторальных сегментах.
Исходно наземные, лишь немногие (Hydrachna/fg, Halacarus/fg) вторично перешли к обитанию в пресной или морской воде. Трахейная система либо отсутствует, либо имеется, причем дыхальца по-разному расположены в разных таксонах. У Prostigmata Kramer 1877 {nom.typ.: Hydrachna/f=Trombidium/g [f:1802; g:1775] (incl. Halacarus)} пара дыхалец находится около хелицер. У Cryptostigmata Canestrini 1891 {«Oribata/fg»} четыре пары дыхалец находятся на вентральной стороне головогруди около оснований ног. У Notostigmata With 1904 {nom.typ.: Opilioacarus/fg [fg:1902]} четыре пары дыхалец находятся на дорсальной стороне первых четырех сегментов опистосомы. У Peritremata Ewing 1909 {nom.typ.: Ixodes/f=Parasitus/g [f:1815; g:1795]} пара дыхалец находится по бокам туловища. У Astigmata Canestrini 1891 {nom.typ.: Acarus/fg (sine Trombidium)} трахейная система отсутствует.
У Prostigmata развитие происходит с особым метаморфозом, при котором чередуются активные и покоящиеся возраста; при этом линька с одного возраста на другой (как с активного на покоящийся, так и с покоящегося на активный) сопровождается дегенерацией конечностей путем антироста и/или отбрасывания с отходами (Jones 1951, 1954). В этом отношении покоящиеся стадии Prostigmata сходны с куколкой насекомых [ср.: Metabola (1)].
Monomerosomata преимущественно мелкие, длина тела 0,1–10 мм. Известны начиная с девона; в современной фауне более 55000 видов (Zhang 2011).
Некоторые авторы считают таксон Monomerosomata полифилетическим.
II. Деление Chelicerata на Xiphosura и Cryptopneustida. Таксон Cryptopneustida Boudreaux 1979a {synn.circ.: Metastomata Weygoldt & Paulus 1979, Sclerophorata Kamenz & Staude & Dunlop 2011} объединяет наземных хелицеровых (Arachnida – см. III) с вымершими морскими Gigantostraca (см. I: 2). Отличатся от мечехвостов следующими аутапоморфиями.
(1) Ротовое отверстие смещено вперед, и в образовании ротового аппарата участвуют не все конечности просомы, а только передние из них: коксы ходильных ног (II–VI посторальных пар) малоподвижно или неподвижно соединены с туловищем, так что по крайней мере коксы V и VI посторальных пар утратили жевательную функцию; по крайней мере задние коксы не сомкнуты медиально, и между ними развит послеротовой стернит – метастома; хилярии полностью утрачены, так что VII посторальный сегмент лишен конечностей.
(2) На опистосоме нет наружных жаберных конечностей.
III. Выделение таксона Arachnida. Традиционно, еще до установления таксона Chelicerata, морских хелицеровых (Xiphosura и Gigantostraca) помещали в таксон Crustacea, а наземных хелицеровых помещали в таксон Tracheata (см. Euarthropoda: «Классификации» I). В соответствии с этой традицией хелицеровых часто делят на морских (Merostoma) и наземных (Arachnida). Таксон Merostoma Thorell & Lindström 1885 объединяет Xiphosura и Gigantostraca, характеризуется лишь первичноморским образом жизни и, судя по всему, является парафилетическим. Таксон Arachnida (паукообразные) рассматривают либо как альтернативный Merostoma, либо как подчиненный в составе Cryptopneustida (см. II).
Arachnida Cuvier 1798 – паукообразные {исходно во французском написании – Arachneides; synn.circ.: Acephala Latreille 1796 = Eleuterosoma Leach 1814 = Octopoda Blainville 1816 (non Octopoda Latreille 1796) = Autarachnae Haeckel 1866 = Tracheopulmonata Lankester 1877 = Chelicerota Lang 1888; nom.typ.: Araneus/fg [fg:(1757)1758]}. Почти идентичные названия Arachnides (во французском и латинском написании) и Arachnidae в начале XIX века употреблялись в трех совершенно разных значениях: (1) для того же таксона, что и Aracnida (Leach 1815b, Blainville 1816, Latreille 1817); (2) для более крупного таксона, включающего всех наземных членистоногих кроме крылатых насекомых (Lamarck 1801, 1815–1822, Latreille 1810) и (3) для более дробного таксона Maripalpi (Latreille 1802, 1806). Для Arachnida указывают следующие апоморфии.
(1) По крайней мере у всех современных представителей фасеточные глаза преобразованы в однолинзовые латеральные глаза (неуникальная апоморфия – см. Указатель признаков [1.20]); помимо латеральных глаз обычно сохраняется пара медиальных глазков [см. Chelicerata (2)]. Однако у некоторых вымерших Toxicura латеральные глаза сохраняют фасеточное строение. У современных Toxicura имеется до пяти пар однолинзовых латеральных глаз и пара глазков; в других таксонах паукообразных число глаз меньше; у Mycetophora и Opiliones глаза утрачены, и имеются только глазки; у Pseudoscorpii, наоборот, глазки утрачены, и имеется только пара однолинзовых глаз; иногда и глаза, и глазки утрачены (табл. IV.3).
![]() Таблица
IV.3. Число пар простых глаз и
глазков в разных таксонах Chelicerophora.
Таблица
IV.3. Число пар простых глаз и
глазков в разных таксонах Chelicerophora.
|
|
oculi |
ocelli |
Максимальный набор глазков (●) и глаз (○ или ░) |
|
Pantropoda |
0 |
≤ 2 |
● ● ● ● |
|
Xiphosura |
facet. |
1 |
░ ● ● ░ |
|
†Gigantostraca |
facet. |
1 |
░ ● ● ░ |
|
†Toxicura-Apoxipodes |
facet. |
1 |
░ ● ● ░ |
|
Toxicura-Dionychopodes |
≤ 5 |
≤ 1 |
○○○○○ ● ● ○○○○○ |
|
Pseudoscorpii |
≤ 2 |
0 |
○○ ◌ ◌ ○○ |
|
Mycetophora |
0 |
1 |
● ● |
|
Anepignathi |
≤ 1 |
0 |
○ ◌ ◌ ○ |
|
Opiliones |
0 |
1 |
● ● |
|
Palpigradi |
0 |
0 |
|
|
Caulogastra |
≤ 3 |
≤ 1 |
○○○ ● ● ○○○ |
|
Monomerosomata |
≤ 2 |
≤ 1 |
○○ ● ● ○○ |
(2) Конечности II–VI посторальных пар [см. Chelicerata (1)] дифференцированы: конечности II пары, называемые педипальпы (pedipalpi), в большинстве случаев специализированы как хватательные или сенсорные, тогда как конечности III–VI пар специализированы как ходильные ноги. Гнатобазы [см. Euarthropoda (3a)] обычно сохраняются только на педипальпах, будучи утрачены на конечностях следующих пар; только у Toxicura и Phalangium/fg1 гнатобазы сохраняются на конечностях следующих двух пар. Среди Arachnida лишь у Palpigradi конечности II пары сходны с ногами следующих пар, но при этом гнатобазы утрачены на всех конечностях.
(3) Претарсус [имеющий двухмыщелковое сочленение – см. Chelicerophora (4)] и развитый на конечностях II–VI или III–VI пар [см. (4)] имеет особое строение. Мышца – сгибатель претарсуса крепится не в тарсусе (который бывает цельным или разделен на проксимальный базитарсус и дистальный телотарсус), а в предыдущем членике или в двух предыдущих члениках – в тибии и в пателле; в то же время мышца – разгибатель претарсуса крепится в тарсусе (если он цельный) или в базитарсусе. Обычно претарсус несет 2 коготка; иногда между ними сохраняется небольшой срединный коготок (у многих Caulogastra и Palpigradi); иногда между коготками имеется непарный мягкий отросток – эмподий (у многих Pseudoscorpii, Mycetophora и Monomerosomata). У большинства Phalangium/fg1 и некоторых других претарсус вторично упрощен и представляет собой один коготок. В остальных отношениях строение конечностей паукообразных разнообразно; общее число члеников [которых исходно 8 – см. Chelicerophora (4)] различно в разных группах, бывает меньше и больше восьми.
В отличие от паукообразных, у Xiphosura и Pantopoda сгибатель и разгибатель претарсуса крепятся в тарсусе, и претарсус представляет собой один коготок. Помимо паукообразных, сгибатель претарсуса крепится в средних члениках ноги у многоножек и насекомых [не имеющих разгибателя – ср.: Atelocerata (3)]. Претарсус с двумя коготками и эмподием, помимо паукообразных, характерен для насекомых [ср.: Hexapoda (6) и Pterygota (5)]. Судя по всему, два коготка на претарсусе независимо возникли у паукообразных и насекомых. Предполагают, что наличие двух коготков связано с адаптацией к передвижению по суше (хотя у многих наземных членистоногих таких коготков нет – см. Указатель признаков [2.58]). У вымерших морских Apoxipodes и Gigantostraca (близких к современным Arachnida-Toxicura) претарсус не имеет двух коготков; предполагают, что это состояние у них первично, следовательно, два коготка независимо возникли в разных группах Arachnida.
(4) Конечности VII пары полностью утрачены (в отличие от Xiphosura).
(5) Все современные паукообразные являются наземными или происходят от наземных предков: большинство из них обитает в воздушной среде, лишь среди клещей (Monomerosomata) имеются таксоны, вторично перешедшие к обитанию в пресной или морской воде (Hydrachna/fg, Halacaris/fg). В связи с наземным образом жизни многие паукообразные имеют мальпигиевы сосуды [см. (5a)] и органы воздушного дыхания в виде книговидных легких [см. (5b)] и/или трахей [см. (5c)]; у некоторых встречаются также втяжные пузырьки [см. (5d)]. Однако, вероятно, разные группы паукообразных независимо перешли к наземному образу жизни (см. «Статус»).
(5 a) Мальпигиевы сосуды паукообразных представляют собой выросты средней кишки и имеют энтодермальное происхождение (в отличие от эктодермальных мальпигиевых сосудов насекомых и многоножек – см. Указатель признаков [4.7.2]). У Phalangium/fg1, Palpigradi и Pseudoscorpii мальпигиевы сосуды отсутствуют. Не исключено, что в разных группах паукообразных мальпигиевы сосуды возникли независимо.
(5 b) У Toxicura и Caulogastra имеются легкие своеобразного строения, называемые «книговидными». Легочный мешок, или атриум легкого, образован впячиванием покрова; внутри он выстлан тонкой кутикулой, а наружу открывается дыхальцем в виде щели, способной закрываться и открываться. Эктодерма, образующая дно этого мешка, утратила строение однослойного эпителия с поляризованными клетками и преобразовалась в особую паренхиматозную ткань, в которой имеются многочисленные плоские клеточные слои, расположенные на равном расстоянии параллельно друг другу и перпендикулярно поверхности. При формировании легкого (происходящего в эмбриогенезе и на ранних стадиях постэмбриогенеза) клетки такого слоя выделяют в обе стороны от себя кутикулу, в результате чего образуют две параллельные кутикулярные пластины, соединенные кутикулярными мостиками (образующимися в зазорах между клетками одного слоя); затем клетки, образовавшие кутикулу, отмирают, а на их месте остаются воздухоносные полости, по толщине равные одной клетке (Farley 2008, 2011). В результате этого параллельные друг другу кутикулярные пластины разделяют правильно чередующиеся воздухоносные полости и кровеносные полости (внутри которых клетки сохраняются). Воздухоносные полости сообщаются с полостью атриума, благодаря чему вся эта ткань обеспечивает газообмен между гемолимфой, поступающей к легкому по кровеносному сосуду [см. Gnathopoda (2)], и воздухом, поступающим в легкое через дыхальце в атриум.
В прошлом была популярной гипотеза о происхождении книговидных легких паукообразных из погрузившихся внутрь тела жабр мечехвостов, которые, в свою очередь, являются видоизмененными конечностями опистосомы [см. Chelicerata (1)] (Lankester 1885). Против этой гипотезы свидетельствует различие в расположении легких и конечностей на сегментах опистосомы у Toxicura и Caulogastra (табл. IV.4). У Toxicura конечности VIII–IX посторальных сегментов (т.е. первых двух сегментов опистосомы) сохранились в виде пары наружных половых крышек и пары гребневидных органов, а на X–XIII посторальных сегментах находятся четыре пары книговидных легких. У Caulogastra, наоборот, на VIII–IX посторальных сегментах находятся две пары книговидных легких, а конечности X–XI посторальных сегментов сохранились в виде наружных паутинных бородавок (имеющихся у пауков). Учитывая, что легкие Toxicura и Caulogastra имеют одинаковое строение и, следовательно, единое происхождение, невозможно представить себе такой путь эволюции, при котором часть конечностей преобразовалась бы в легкие, а часть сохранились бы в виде наружных придатков взаимоисключающим способом у Toxicura и Caulogastra. Книговидные легкие паукообразных принципиально отличаются от жаберных конечностей тем, что у книговидных легких воздухоносные каналы образуются на месте отмерших клеток – т.е. в исходно внутренней среде организма, а у конечностей полости и каналы могут образовываться только как впячивания покровов – т.е. в исходно наружной среде. Так что эти образования не гомологичны.
Можно предположить, что книговидные легкие либо возникли у общего предка Arachnida, но вторично утратились у всех паукообразных, кроме Toxicura и Caulogastra; либо возникли только у общего предка Pulmonariae (см. IV).
![]() Таблица
IV.4. Расположение пар конечностей и пар легких на посторальных сегментах у
мечехвостов, скорпионов и
пауков
Таблица
IV.4. Расположение пар конечностей и пар легких на посторальных сегментах у
мечехвостов, скорпионов и
пауков
|
Тагмы: |
просома | опистосома | |||||||||||||||||
| Постоальные сегменты: | I | II |
III |
IV |
V |
VI |
VII |
VIII |
IX |
X |
XI |
XII |
XIII |
XIV |
XV | XVI | XVII | XVIII | XIX |
|
Xiphosura: конечности |
chel |
p |
p |
p |
p |
p |
chi |
go |
br |
br |
br |
br |
br |
– |
|||||
|
Toxicura: конечности легкие |
chel |
pp |
p |
p |
p |
p |
– |
go |
pect |
– + |
– + |
– + |
– + |
– |
M | M | M | M | M |
|
Caulogastra: конечности легкие |
chel |
pp |
p |
p |
p |
p |
– |
– + |
– + |
w |
w |
– |
– |
– |
– | – | – | – | |
Условные обозначаения: br – жабры; chel – хелицеры; chi – хилярии; M – сегменты метасомы; pp – педипальпы; p – ходильные ноги I–IV пар; go – генитальные крышки; pect – гребневидные органы; w – паутинные бородавки; + – книговидные легкие.
(5 c) Многие Arachnida дышат трахеями. При этом в разных таксонах трахеостии открываются на разных сегментах опистосомы или просомы (см. I: Caulogastra, Pseudoscorpii, Mycetophora, Phalangium/fg1 и Monomerosomata). Это свидетельствует о том, что трахейная система арахнид возникала несколько раз независимо, так же как она независимо многократно возникала в других группах наземных членистоногих [см. Gnathopoda (3c)].
(5 d) У некоторых представителей Monomerosomata на коксах второй пары ног личинки и на половых крышках, образованных конечностями I–III сегментов опистосомы (т.е. VIII–X посторальных сегментов) имеются парные втяжные пузырьки; подобные пузырьки имеются также у Palpigradi на I–V сегментах опистосомы (т.е. на VIII–XII посторальных сегментах) (Захваткин 1952). Подобные коксальные пузырьки имеются у некоторых других наземных членистоногих – у некоторых онихофор и различных Atelocerata (см. Указатель признаков [3.36]).
Статус Arachnida. По мнению некоторых авторов, таксон Arachnida, объединяющий наземных хелицеровых, является полифилетическим. В пользу этого говорит то, что относящиеся к Arachnida современные наземные скорпионы (Dionychopodes) имеют синапоморфии с вымершими морскими Apoxipodes (см. I: 3 – Toxicura); некоторые из этих синапоморфий являются также общими для Gigantostraca. Gigantostraca и Toxicura обладают единой специализацией туловищных сегментов: просома слитная, а опистосома подразделена на более широкую 7-сегментную мезосому (заканчивающуюся XIV сегментом) и более узкую 5-сегментную метасому (начинающуюся с XV сегмента). У других хелицеровых этой особенности нет: у Pseudoscorpii, Mycetophora, Phalangium/fg1, Palpigradi, Caulogastra и Monomerosomata сегменты до XIV и после XV принципиально не различаются; для Xiphosura этот признак неизвестен, поскольку у них нет XV–XIX сегментов. На основе наличия такой синапоморфии у Gigantostraca и Toxicura делают вывод, что разные группы хелицеровых переходили к жизни на суше независимо несколько раз (по крайней мере не менее двух раз): наземные скорпионы (Toxicura-Dionychopodes) произошли непосредственно из палеозойских морских предков – Toxicura-Apoxipodes и Gigantostraca, тогда как другие наземные паукообразные произошли из каких-то иных, неизвестных нам, морских предков.
IV. Деление Arachnida на легочных и безлегочных (табл. IV.5).
Таксон Pulmonariae Latreille 1817 {synn.circ.: Pulmonaria Westwood 1838 = Aerobranchia Lankester 1881 = Embolobranchia Lankester 1888} объединяет Toxicura и Caulogastra. Он характеризуется наличием на опистосоме своеобразных книговидных легких – по паре на первых двух сегментах опистосомы у Caulogastra и по паре на следующих четырех сегментах у Toxicura [см. III: Arachnida (5b)]. У некоторых Caulogastra утрачена вторая пара или обе пары легких; в дополнение к легким могут быть развиты трахеи.
Таксон Lipobranchia Lankester 1881 (non Lipobranchia Haeckel 1866) {syn.circ.: Trachearia Westwood 1838 (non Tracheariae Latreille 1817–1825)}. Объединяет всех прочих паукообразных – Pseudoscorpii, Mycetophora, Phalangium/fg1 и Monomerosomata. Характеризуется отсутствием книговидных легких. Органами дыхания служат трахеи, которые, вероятно, возникали несколько раз независимо, поскольку их устья расположены на разных сегментах [см. III: Aracnida (5c)]. Разные авторы считают этот таксон либо полифилетическим, либо голофилетическим. Разногласия вызывает также положение Palpigradi, которые из-за своих мелких размеров утратили органы дыхания: таксон Apulmonata Firstman 1973 (= Lipobranchia s.l.) включает Palpigradi, а таксон Holotracheata Börner 1902 (= Lipobranchia s.str.) не включает Palpigradi.
Делению Arachnida на Pulmonariae и Lipobranchia противоречит тот факт, что по строению педипальп Toxicura и Pseudoscorpii имеют глубокое сходство (см. VI).
V. Выделение таксона Lipoctena (табл. IV.5). Всех паукообразных кроме Toxicura иногда объединяют в таксон Lipoctena Pocock 1893c {syn.circ.: Epectinata Lankester 1910}. Если исходить из предположения, что Arachnida – голофилетический таксон, вышедший на сушу один раз (см. III) и учесть, что относящееся к нему Toxicura имеют непосредственное родство с первичноморскими Apoxipodes и более дальнее родство с первичноморскими Gigantostraca, получается вывод, что все Lipoctena произошли непосредственно от Toxicura. В таком случае аутапоморфиями Lipoctena являются утрата тех структур, которые уникальны для Toxicura – гребневидных органов, разделения брюшка на мезосому и тонкую пятисегментную метасому и др. У Lipoctena рабдомеры глаз имеют сетевидное расположение, тогда как у Toxicura они сохраняют звездчатое расположение, унаследованное от предковых фасеточных глаз (Weygoldt & Paulus 1978).
VI. Объединение Toxicura и Pseudoscorpii (табл. IV.5). У скорпионов (Toxicura) и ложноскорпионов (Pseudoscorpii) педипальпы преобразованы в очень крупные клешни, имеющие уникальное строение. Педипальпа состоит из 6 члеников: 1-й и 2-й членики короткие, 3-й членик длинный и сгибается назад, 4-й членик сгибается вперед, 5-й членик сильно утолщен и несет неподвижный палец клешни, 6-й членик (тарсус) причленен вентральнее неподвижного пальца и представляет собой подвижный палец клешни; 7-й членик (претарсус) полностью утрачен. Как и на других ногах, тарсус двигается только мышцами-сгибателями, идущими в два предыдущих членика, и не имеет разгибателей; так что к основанию подвижного пальца подходят две мышцы: одна, очень мощная, крепится в утолщенной части 5-го членика и обеспечивает сильное сжатие клешни; вторая, тонкая, тянется из 4-го членика, изгибаясь и проходя через узкое подвижное сочленение между четвертым и пятым члениками, так что она, вероятно, не способна к каким-либо действиям. В результате клешни скорпионов и ложноскорпионов способны только сжиматься с большой силой, используя лишь одну мышцу из двух имеющихся; разжимание клешни происходит гидравлическим способом. Такое нерациональное строение клешней педипальп скорпионов и ложноскорпионов принципиально отличается от строения других клешней. У Chelicerata и Eucrustacea, в отличие от Atelocerata, претарсус исходно двигается сгибателем и разгибателем, крепящимися в предыдущем членике (или в двух предыдущих члениках); способность претарсуса не только к сгибанию, но и к активному разгибанию явилась предпосылкой к преобразованию некоторых конечностей в клешни, где претарсус образует подвижный палец: такие клешни независимо возникли на хелицерах, на всех ногах у Xiphosura и на различных конечностях некоторых Eucrustacea. У Atelocerata клешни почти не встречаются, поскольку у них претарсус исходно не имеет разгибателя и, если он образует подвижный палец клешни, такая клешня не может активно разжиматься. На педипальпах скорпионов и ложноскорпионов сначала утратился претарсус, который мог бы быть эффективным подвижным пальцем клешни, а потом образовалась клешня, у которой подвижным пальцем служит наименее пригодный для этой функции тарсус. Такое одинаковое нерациональное строение клешней является серьезным доводом в пользу сестринского родства Toxicura и Pseudoscorpii.
VII. Объединение Mycetophora и Pseudoscorpii (табл. IV.5). Таксон Haplocnemata Börner 1904b {synn.circ.: Pseudoscorpiones Latreille 1825 = Epatellata Lameere 1933 = Apatellata Hammen 1977 = Chelonetomorphae Savory 1977} объединяет сольпуг (Mycetophora) и ложноскорпионов (Pseudoscorpii) и характеризуется тем, что хелицеры [исходно трехчлениковые клешневидные – см. Chelicerophora (2)] утратили проксимальный членик, так что стали двучлениковыми и направлены вершиной вперед; подвижный палец причленен не дорсально (как у большинства Chelicerophora), а вентрально. Представление о голофилии Haplocnemata противоречит представлению о голофилии таксона, объединяющего Pseudoscorpii и Toxicura.
Таблица IV.5 (RTF)
* * *