КЛАДОЭНДЕЗИС НАСЕКОМЫХ

![]()
|
КЛАДОЭНДЕЗИС НАСЕКОМЫХ |
|||||||
|
|
|
fg | |||||
Глава VI. Систематика Hexapoda: общая классификация и первичнобескрылые
VI-1. Hexapoda Latreille 1802 – шестиножки, или насекомые
(рис. 6.1.1–10.16.2)
|
Название, однозначно соответствующее по циркумскрипции: — Hexapoda Latreille 1802a (T.2: 344-345) (non Hexapoda Batsch 1789; nec Hexapoda Latreille 1796). По циркумскрипции также соответствует: — «Insecta» sensu Leach 1815a (non Linnaeus 1758). Иерархическое название: Pediculus/f3=Scarabaeus/g3 [f:1777; g:L.1758(gen.170)] (sine Scolopendra; incl. Podura).
|
|
Ранг. От подкласса до подтипа, обычно класс или надкласс.
Названия. Часто для этого таксона используют название Insecta; это же название используют и для многих других таксонов (см. I.6.6.2). В отличие от названия Insecta, другое широко признанное название – Hexapoda, всегда используется только для таксона одной и той же циркумскрипции. Исходно (Latreille 1802) таксон Hexapoda был лишь упомянут в обсуждении, но не использован в основной классификации; в последующем (Blainville 1816) таксон Hexapoda стал общепризнанным. Лишь в конце XVIII и начале IX века некоторые авторы использовали название Hexapoda в двух других значениях, не связанных с вышеназванным: (1) для тех шестиногих насекомых, которых ранее помещали в отряд Aptera Linnaeus 1758, был предложен таксон Hexapoda Batsch 1789, объединявший первичнобескрылых насекомых с вторичнобескрылыми вшами и блохами; (2) для паразитических бескрылых мух Nycteribia и шестиногих личинок клещей (Monomerosomata в составе Chelicerata) ошибочно выделялся таксон, называвшийся Hexapoda Latreille 1796.
Аутапоморфии Hexapoda. Общая характеристика Hexapoda хорошо известна, поскольку она повторяется в многочисленных учебниках и пособиях, изданных в разных странах начиная с середины XIX века и по сей день. Однако из большинства этих источников неясно, какие именно признаки являются аутапоморфиями Hexapoda, а какие принадлежат вышестоящим или подчиненным таксонам. Поэтому у некоторых людей даже сложилось впечатление, что у Hexapoda нет аутапоморфий, и что этот таксон может быть не голофилетическим, а полифилетическим или парафилетическим (см. «Статус»). В действительности уникальные аутапоморфии Hexapoda содержатся среди общеизвестных признаков строения ротового аппарата [см. (2) и (3)], груди [см. (4)], ног [см. (5) и (6)], брюшка [см. (7) и (8)] и др. [см. (9) и (10)].
(1) Постоянное число сегментов и эпиморфоз. Число посторальных сегментов постоянно и равно 17; из них первые 4 сегмента входят в состав головы, следующие 3 сегмента образуют грудь, и следующие 10 сегментов образуют брюшко [исключение составляют ногохвостки и бессяжковые, у которых число сегментов брюшка уменьшено – см. ниже, Ellipura (3)]; все 17 сегментов формируется при эмбриональном развитии под яйцевыми оболочками и их число остается неизменным в процессе постэмбриогенеза, т.е. имеет место эпиморфоз. Число обособленных ганглиев вентральной нервной цепочки [которое исходно соответствует числу сегментов – см. Metameria (2)] у насекомых всегда меньше семнадцати: помимо того, что первый посторальный ганглий (тритоцеребрум) вошел в состав надглоточного ганглия [см. Euarthropoda (6)], а II–IV посторальные ганглии, иннервирующие ротовые придатки головы, слиты в подглоточный синганглий, из десяти сегментов брюшка только первые 8 имеют собственные ганглии, тогда как IX и X сегменты брюшка иннервируются от ганглия VIII сегмента брюшка. Во многих таксонах насекомых произошло дальнейшее уменьшение числа ганглиев за счет слияний тех или иных ганглиев груди и/или брюшка. В ходе постэмбриогенеза число обособленных ганглиев либо остается постоянным, либо меняется при неизменном числе сегментов.
Вероятно, исходным типом развития у членистоногих был анаморфоз – развитие с увеличением числа сегментов в эмбриогенезе, свойственное многим Eucrustacea и Myriapoda [см. Metameria (1b)]. Высказывалось представление о том, что среди Hexapoda первичный анаморфоз сохранился у Protura; однако здесь речь идет о вторичном явлении [см. ниже, Protura (6)]. У некоторых Pterygota с усложненным метаморфозом личинки имеют уменьшенное число сегментов брюшка; такие случаи увеличения числа сегментов в постэмбриогенезе являются вторичными (см. Указатель признаков [3.2]). Независимо от Hexapoda эпиморфоз возник у Chilopoda-Epimorpha и у некоторых других членистоногих (см. Указатель признаков [5.6]).
Высказывались утверждения о наличии у Hexapoda большего, чем семнадцать, числа исходных сегментов. В составе головы некоторые авторы усматривают более четырех исходных сегментов, предполагая, что исходно существовали отдельные сегменты, несущие антенны, глаза и некоторые другие детали строения головы. Однако антенны и глаза относятся не к посторальным сегментам, а к преоральной области тела [см. Metameria (2)]. Была высказана гипотеза о происхождении трех сегментов груди из исходных шести сегментов [см. (4c)]. Распространено ошибочное утверждение, что в составе брюшка, помимо десяти сегментов, имеются вестигии одиннадцатого сегмента [см. (8b)].
(1 a) Дифференциация сегментов и их конечностей. Преоральная область и 17 посторальных сегментов, составляющих тело насекомого, дифференцированы следующим образом.
Голова (caput) у Hexapoda состоит из преорального отдела, несущего антенны [см. Euarthropoda (6)], фасеточные глаза и глазки [см. Euarthropoda (4)], и четырех посторальных сегментов (I–IV сегменты). Поскольку I посторальный сегмент лишен конечностей [см. Atelocerata (1)], голова несет 3 пары посторальных конечностей, служащих ротовыми придатками – мандибулы, максиллы и нижнюю губу. Исходное положение головы гипогнатное, т.е. направленное ротовыми придатками вентрально, так что мандибулы расположены впереди максилл, а максиллы – впереди нижней губы, в соответствии с исходным расположением их сегментов; в ходе эволюции положение головы многократно менялось на прогнатное (т.е. ротовыми придатками вперед, так что мандибулы расположены дорсально, а нижняя губа вентрально) и обратно на гипогнатное. Ротовой аппарат Hexapoda состоит из непарной верхней губы [см. Euarthropoda (7)], пары одночлениковых мандибул [см. Mandibulata (1) и Atelocerata (4)], пары максилл сложного строения [см. (2)], пары суперлингв [см. Mandibulata (3)], гипофаринкса [см. Mandibulata (2)] и нижней губы сложного строения [см. (3)]. Исходно ротовой аппарат грызущий: мандибулы служат для разгрызания пищи, максиллы манипулируют частицами пищи и перемещают их в рот, нижняя губа удерживает пищу снизу, а телоподиты максилл и нижней губы служат осязательными органами – щупиками. Исходной пищей насекомых являются детрит, грибы, водоросли, споры и пыльца растений. В некоторых таксонах Hexapoda ротовой аппарат преобразовался из грызущего в иной – сосущий, колюще-сосущий, лижуще-сосущий, фильтрующий, скребущий и др. (см. Указатель признаков [1.36]), причем в каждом случае произошли уникальные преобразования или утрата определенных частей ротового аппарата. Некоторые насекомые приобрели ту или иную пищевую специализацию – хищничество, фитофагию или иную. В большинстве случаев нет однозначной связи между модификацией ротового аппарата и пищевой специализацией. В строении ротового аппарата насекомых, помимо признаков, унаследованных от предков Hexapoda, и признаков, приобретенных в тех или иных частных таксонах насекомых, присутствуют аутапоморфии Hexapoda – это исходный для Hexapoda план строения максилл [см. (2)] и исходный для Hexapoda план строения нижней губы [см. (3)].
Грудь, или торакс (thorax), Hexapoda состоит из трех сегментов (V–VII посторальные сегменты), несущих по паре ног. Помимо Hexapoda, отличие тех же трех сегментов от последующих сегментов туловища имеет место у Progoneata, но особенности строения трех сегментов груди, свойственные Hexapoda, являются уникальными и не встречаются у других животных [см. (4)]. Членики ног у любых членистоногих принято называть терминами, исходно введенными для Hexapoda – кокса, трохантер, фемур, тибиа, тарсус и претарсус; благодаря этому может сложиться впечатление, что ноги всех членистоногих имеют единый план строения. В действительности лишь некоторые членики ног гомологичны у разных членистоногих, а строение ног Hexapoda является уникальным [см. (5) и (6)].
Брюшко, или абдомен (abdomen) у различных Hexapoda настолько разнообразно, что до недавнего времени не было ясного представления о числе входящих в него сегментов и об их исходном строении. Критическое сопоставление имеющихся данных и собственные исследования позволили автору этой книги сформулировать новую общую характеристику брюшка Hexapoda и выявить аутапоморфии Hexapoda в строении брюшка. Брюшко гексапод состоит из 10 сегментов, являющихся VIII–XVII посторальными сегментами тела. Первые 9 из этих сегментов имеют повторяющиеся черты строения [см. (7)], а десятый сегмент брюшка имеет особое строение [см. (8)]. У многих насекомых число сегментов брюшка кажется меньшим десяти, так как последние сегменты бывают сильно модифицированы, уменьшены и/или втянуты в предыдущие; лишь у ногохвосток и протур число сегментов брюшка действительно меньше десяти [см. ниже, Ellipura (3)].
Синонимом термина «абдомен» является термин «уросома» (urosoma); для краткости сегменты брюшка насекомых называют уромерами (uromeri, или uromera), а их тергиты и стерниты, соответственно, уротергитами и уростернитами.
(1 b) Эволюционная корреляция сегментов. Эволюционная корреляция сегментов в некоторых случаях совпадает с их дифференциацией, а в некоторых случаях с ней не совпадает.
Сегменты, входящие с в состав головы, исходно не имеют эволюционной корреляции: их придатки – мандибулы, максиллы и нижняя губа имеют принципиально разное строение и в ходе эволюции не приобретают общих черт. Однако несмотря на принципиальное различие между строением максилл и нижней губы, в отдельных таксонах насекомых максиллярные щупики [исходно 5-члениковые – см. (2c)] и лабиальные щупики [исходно 3-члениковые – см. (3с)] приобретают эволюционную корреляцию (корреляция 6 на табл. VI.1): несмотря на различное число члеников, дистальные членики лабиального и максиллярного щупиков могут иметь общие особенности, специфичные для таксона.
Три сегмента груди исходно имеют эволюционную корреляцию (корреляция 1 на табл. VI.1), выражающуюся в повторяющихся чертах строения всех трех пар ног [см. (5) и (6)] и в сходном расположении туловищных швов и аподем [см. Pterygota (3)]. Кроме того, в строении второго и третьего сегментов груди во многих случаях проявляется особая эволюционная корреляция, не распространяющаяся на строение переднегруди (корреляция 3 на табл. VI.1): у крылатых насекомых средне- и заднегрудь несут крылья и имеют особые черты строения, образуя птероторакс [см. ниже, Pterygota (4)]; у Microcoryphia ноги этих же сегментов несут стилусоподобные придатки, отсутствующие на передних ногах [см. ниже, Microcoryphia (6)]; в исходном плане строения Hexapoda сегментарные грудные дыхальца имеются только на средне- и заднегруди [см. (4b)].
Из десяти сегментов брюшка исходно эволюционно коррелированы только девять сегментов, с первого по девятый [см. (7)] (корреляция 2 на табл. VI.1). У многих насекомых сквозная корреляция всех девяти сегментов брюшка утрачена, но сохраняется корреляция между некоторыми из этих сегментов.
В некоторых случаях наблюдается эволюционная корреляция между среднегрудью, заднегрудью и первыми девятью сегментами брюшка (т.е. между всеми сегментами туловища кроме первого и последнего) (корреляция 4 на табл. VI.1).
Такая корреляция может проявляться в строении отростков и вторичных придатков туловища у личинок некоторых Metabola (рис. 10.10.2B). В частности, у личинок некоторых представителей таксона Metriorrhynchus/fg (относящегося к жукам-краснокрылам – Lycus/fg) на боках этих сегментов имеются подвижные придатки, сериально гомологичные урогомфам [придаткам IX сегмента личинок жуков – см. Elytrophora (12)]; у личинки на рис. 10.2.5D одновременно проявляется и эволюционная корреляция 4 (в форме латеральных придатков), и корреляция 2 (в форме парных дорсальных придатков), и корреляция 5 (в парном строении всех туловищных тергитов), и корреляция 3 (в сходной форме склеритов птероторакса). У личинок некоторых Metabola на сегментах туловища имеются метамерные склеротизованные выпуклости, которые на средне- и заднегруди являются предшественниками куколочных протоптеронов (рис. 10.7.3A,C). Такая же корреляция может проявляться в строении некоторых других туловищных образований, например, трахейных жабр (рис. 10.15.1C).
Возможно, у предка Pterygota на туловищных сегментах имелись парные подвижные придатки с такой же эволюционной корреляцией, и в дальнейшем они дали начало крыльям на средне- и заднегруди и тергалиям на I–IX сегментах брюшка [см. Pterygota (4D)].
![]() Табл. VI.1. Типы эволюционной корреляции, встречающиеся у Hexapoda
Табл. VI.1. Типы эволюционной корреляции, встречающиеся у Hexapoda
| Посторальные сегменты: | I | II | III | IV | V | VI | VII | VIII | IX | X | XI | XII | XIII | XIV | XV | XVI | XVII | |
| Специализация сегментов: | – | md | mx | lb | prth | msth | mtth | urI | urII | urIII | urIV | urV | urVI | urVII | urVIII | urIX | urX | |
| Тагмы: | голова | грудь | брюшко | |||||||||||||||
| Случаи эволюционной корреляции: |
1 | строение ног | ||||||||||||||||
| 2 | строение уромеров; стерностили и кинетапофизы |
|||||||||||||||||
| 3 | птероторакс, крылья |
|||||||||||||||||
| 4 | некоторые придатки туловища | |||||||||||||||||
| 5 | форма туловищных тергитов | |||||||||||||||||
| 6 | форма щупиков |
|||||||||||||||||
(2) Максиллы. В ротовом аппарате, помимо мандибул [специализированных жующих конечностей II посторальной пары – см. Mandibulata (1)] имеется еще одна пара подвижных специализированных жующих конечностей – нижних челюстей, или максилл (maxillae), соответствующих максиллам первым у Eucrustacea, т.е. конечностям III посторальной пары, вовлеченным в ротовой аппарат [см. Mandibulata (2)]. Строение максилл Hexapoda является уникальным и едино как для Entognatha (рис. 6.3.1E), так и для Amyocerata (рис. 6.7.1D, 9.1.3F).
(2 a) Строение максиллы (рис. 6.7.1D, 9.1.3F). Проксимальная часть максиллы, входящая в состав стенки головы, разделена на две подвижно сочлененные части – поперечно направленный кардо (cardo) и продольно направленный стипес (stipes); каждая из этих частей не является самостоятельным члеником и непосредственно причленена к голове своей длинной медиальной стороной; от заднего (вентрального) края каждой из них идут мышцы к эндоскелету головы – к фультуре [см. ниже, Entognatha (1)] или к тенториуму [см. ниже, Amyocerata (3)]; в случаях утраты тенториума эти мышцы крепятся непосредственно к головной капсуле.
Выступающими частями максиллы являются лациния, галеа и щупик. На дистальном конце стипес несет жевательную долю – лацинию (lacinia), от основания которой отходят две мышцы, идущие впереди (дорсальнее) мышц стипеса и кардо: одна из мышц лацинии идет в стипес, другая – к головной капсуле. Исходно лациния склеротизована, на вершине заострена или зазубрена (обычно заканчивается двумя или тремя зубцами, направленными дистально-медиально); проксимальнее вершинных зубцов ее медиальный край несет зубцы и/или крепкие щетинки. Латеральнее и дорсальнее лацинии от стипеса отходит другой придаток – галеа (galea); от ее основания идет мышца в стипес, эта мышца лежит позади (вентральнее) всех прочих мышц стипеса. Исходно галеа не склеротизована, туповершинная. Латеральнее и дорсальнее галеи от стипеса отходит максиллярный щупик (palpus maxillaris), от основания которого мышцы идут к заднему (вентральному) краю стипеса. Исходно щупик расчленен, и в каждом его членике кроме последнего имеется одна или несколько мышц, двигающих следующий членик [см. (2c)].
Благодаря тому, что проксимальная часть максиллы имеет два подвижно сочлененных склерита (кардо и стипес), к каждому из которых подходят мышцы, максилла способна совершать сложные движения, двигаясь вперед-назад и медиально-латерально; в результате этого лацинии максилл, вооруженные зубцами и щетинками, способны манипулировать частицами пищи и загребать их в рот.
У некоторых Hexapoda максиллы модифицированы так, что утратили те или иные части, иногда максиллы совсем утрачены (см. Указатель признаков [1.55–1.60]).
У Entognatha и Triplura максиллы сохраняют кардо, стипес, лацинию, галею и щупик. У личинок поденок максиллы сохраняют кардо, стипес, лацинию и, как правило, щупик, но утратили обособленную галею [см. ниже, Ephemeroptera (5)]. У стрекоз максилла утратила либо галею, либо щупик [см. ниже, Odonata (3b)]. У Idioprothoraca, Rhipineoptera и Zoraptera максиллы сохраняют кардо, стипес, лацинию, галею и щупик [о лацинии Zoraptera – см. Parametabola (1)]. У Acercaria максиллы утратили кардо и подвижную галею, а стипес и лациния сильно преобразованы; щупик сохраняется или утрачен [см. ниже, Acercaria (1), Parapsocida (2), Eupsocida (2), Parasita (5), Condylognatha (1b), Thysanoptera (2), Arthroidignatha (1b)]. У большинства Eleuterata максиллы сохраняют кардо, стипес, лацинию, галею и щупик. У Strepsiptera максиллы вестигиальные, как и весь ротовой аппарат [см. ниже, Strepsiptera (1), (2), (3)]. У имаго всех Neuropteroidea и у личинок Rhaphidioptera и Meganeuroptera максиллы сохраняют кардо, стипес, лацинию, галею и щупик; при этом у личинок вислокрылок максиллы по-разному модифицированы [см. ниже, Nothomegaloptera (1) и Eumegaloptera (1)]. У личинок Birostrata максиллы, вошедшие в состав парного колюще-сосущего ротового аппарата, уникальным образом модифицированы [см. ниже, Birostrata (1)]. У Hymenoptera максиллы сохраняют кардо, стипес, лацинию, галею и щупик, бывают по-разному модифицированы у личинок и имаго. Среди Enteracantha, у личинок и у имаго Scorpiomuscae, Nannomecoptera, Metamecoptera и Raphioptera максиллы сохраняют кардо, стипес, лацинию, галею и щупик; у имаго блох максиллы сильно модифицированы, без кардо и галеи, с погруженным стипесом, колющими лациниями и модифицированным щупиком [см. ниже, Aphaniptera (1b)]. У личинок Diptera максиллы либо сохраняют кардо, стипес, лацинию, галею и щупик, либо имеют упрощенное строение; у имаго двукрылых галеа утрачена, а максиллы исходно специализированы как колюще-режущие [см. ниже, Diptera (1Ad)]; у имаго Cyclorapha и некоторых других максиллы сохранили только щупики, тогда как их остальные части вошли в состав непарного хоботка [см. ниже, Cyclorapha (2)]. Среди Amphiesmenoptera, у личинок и у имаго Trichoptera и Protolepidoptera максиллы сохраняют кардо, стипес, лацинию, галею и щупик; у имаго Glossolepidoptera максиллы утратили лацинии, а их галеи преобразовались в сосущий хоботок [см. ниже, Glossolepidoptera (1)].
(2 b) Сравнение. По своему происхождению кардо, стипес и лациния, вероятно, соответствуют коксоподиту, а щупик – телоподиту. Происхождение галеи неясно: это либо придаток коксоподита, либо внутренняя ветвь телоподита, соответствующая эндоподиту ракообразных [ср.: Eucrustacea (2)]. Если галеа – это эндоподит, то щупик представляет собой не весь телоподит, а лишь его наружную ветвь – экзоподит, сохраняющий членистое строение (в отличие от нечленистого максиллярного экзоподита у Eucrustacea).
Строение максилл Hexapoda уникально. У различных Eucrustacea строение максилл I разнообразно, но не бывает таким, как у Hexapoda. Среди Myriapoda настоящие максиллы (т.е. жевательные конечности III посторальной пары) имеются только у Symphyla; в отличие от максиллы Hexapoda, максилла Symphyla нерасчлененная и оканчивается лишь одним жевательным выступом [ср.: Symphyla (3) и рис. 5.2.1B] (неясно, является ли это первичным состоянием или вторичным упрощением, подобно вторичному упрощению максилл у некоторых Hexapoda).
Возможно, сложное строение максилл Hexapoda отчасти унаследовано ими от общего предка Mandibulata, так что апоморфиями Hexapoda являются не все, а лишь некоторые из названных черт строения максиллы.
(2 c) Число члеников максиллярного щупика. По-видимому, исходным для Hexapoda является наличие 5 члеников в максиллярном щупике [в отличие от 3-членикового лабиального щупика – см. (3c)]. Иногда число члеников уменьшено благодаря редукции или слиянию тех или иных члеников, иногда щупики утрачены; реже число члеников щупика увеличено благодаря вторичному расчленению некоторых члеников (см. Указатель признаков [1.61]: табл. C). У всех Entognatha максиллярные щупики в той или иной мере уменьшены и упрощены в связи с тем, что максиллы погружены в карманы (рис. 6.3.1E); таким образом, неясно, являются ли пятичлениковые максиллярные щупики исходно свойственными Hexapoda или лишь Amyocerata.
У Zygentoma максиллярные щупики 5-члениковые. У Microcoryphia максиллярные щупики сильно удлинены и число их члеников увеличено до 7 [см. ниже, Microcoryphia (2)]. У поденок и стрекоз число члеников максиллярного щупика различным образом сокращено [см. ниже, Ephemeroptera (6) и Odonata (3b)]. У всех Idioprothoraca и большинства Rhipineoptera максиллярные щупики 5-члениковые (в единичных случаях 4-члениковые) [см. ниже, Idioprothoraca (6) и Rhipineoptera (3)]. Среди Parametabola 5-члениковые максиллярные щупики сохраняются у Zoraptera. У Acercaria максиллярные щупики имеют сокращенное число члеников: имеется не более 4 члеников, лишь у некоторых трипсов щупики вторично расчленены [см. ниже, Acercaria (2)].
У многих Metabola по крайней мере на стадии имаго максиллярные щупики 5-члениковые; у личинок Metabola максиллярные щупики обычно также 5-члениковые, но значительно меньшего размера, с сильно укороченными члениками. У имаго и личинок жуков число члеников максиллярного щупика, вероятно, исходно уменьшено до 4, иногда число члеников сокращено до 3 или 2, реже вторично увеличено до 5. У имаго и личинок веерокрылых максиллы сильно видоизменены [см. ниже, Strepsiptera (2)]. У имаго сетчатокрылообразных максиллярные щупики обычно 5-члениковые, лишь у некоторых Eumegaloptera 4-члениковые; у личинок Birostrata максиллярный щупик не расчленен и уникальным образом модифицирован [см. ниже, Birostrata (1)]; у личинок других сетчатокрылообразных (Rhaphidioptera и Meganeuroptera) максиллярный щупик имеет то же число члеников, что у имаго [см. ниже, Rhaphidioptera (1), Nothomegaloptera (1) и Eumegaloptera (1)]. У имаго перепончатокрылых число члеников максиллярного щупика, вероятно, исходно увеличено до 6; бывает от 1 до 6 члеников; у личинок перепончатокрылых максиллярный щупик 5-члениковый или с меньшим числом члеников [см. ниже, Hymenoptera (1)]. У имаго всех Enteracantha (скорпионницы и блохи) максиллярные щупики 5-члениковые [у блох кажутся 4-члениковыми – см. ниже, Aphaniptera (1b)]; у личинок число члеников максиллярного щупика уменьшено до 3 (у большинства скорпионниц) или до 2 (у ледничников и блох) [см. ниже, Calyptroptera (7)]. У имаго Diptera максиллярные щупики исходно 5-члениковые, в некоторых таксонах число члеников сокращено; в частности, у Brachocera число члеников сокращено до двух, у Brachocera-Cyclorapha – до одного [см. ниже, Brachocera (2) и Cyclorapha (2)]; у личинок двукрылых максиллярные щупики обычно нерасчлененные. У имаго Trichoptera максиллярные щупики исходно 5-члениковые; у некоторых представителей модифицированы, с уменьшенным числом члеников; иногда вестигиальные или утрачены; у личинок Trichoptera максиллярные щупики 4-члениковые. У имаго Lepidoptera максиллярные щупики исходно 5-члениковые, но у большинства представителей вестигиальные, с уменьшенным числом члеников, или утрачены [см. ниже, Lepidoptera (3)]; у личинок Lepidoptera максиллярные щупики имеют не более 3 члеников.
Некоторые авторы считают определенные членики максиллярного щупика Hexapoda гомологичными вертлугу, бедру, голени и лапке ноги (Snodgrass 1935, Grasse 1949); такая гомологизация крайне сомнительна, так как максиллы Mandibulata специализировались в качестве сугубо ротовых придатков раньше, чем возникло характерное для Hexapoda расчленение ноги [см. (5)], а эволюционная корреляция между строением ноги и строением максиллярного щупика отсутствует. Исходно нет также эволюционной корреляции в строении максиллярного и лабиального щупиков, но иногда такая корреляция возникает вторично [см. (1b)].
(3) Нижняя губа. Конечности IV посторальной пары [соответствующие максиллам II многих Eucrustacea – см. Mandibulata (2)] слились медиально своими коксоподитами, образовав непарную нижнюю губу, или лабиум (labium) (рис. 7.2.3H); нижняя губа является третьей парой ротовых конечностей [после мандибул и максилл – см. (2)] и последней парой конечностей головы [см. (11)]. Гипофаринкс [непарный выступ между основаниями максилл – см. Mandibulata (2)] оказался расположенным между ротовым отверстием и нижней губой. Протоки, идущие от пары желез, принадлежащих IV посторальному сегменту, обычно соединяются в непарный проток, который открывается непарным отверстием у основания нижней губы, обычно между нижней губой и гипофаринксом; эти железы называют лабиальными (glandulae labiales), или слюнными (g. salivares).
(3 a) Строение нижней губы. Нижняя губа дистально несет две пары отчлененных придатков: более медиальные придатки – глоссы (glossae) и более латеральные придатки – параглоссы (paraglossae); латеральнее параглосс причленяется пара членистых лабиальных (нижнегубных) щупиков (palpi labiales). К основанию каждой глоссы, параглоссы и щупика подходит одна или несколько мышц, неподвижные концы которых крепятся в непарной части нижней губы. Внутри глосс и параглосс мышц нет. Щупик обычно расчленен, и в каждом его членике кроме последнего имеется одна или несколько мышц, двигающих следующий членик [см. (3c)]. У некоторых Hexapoda глоссы и/или параглоссы слиты или редуцированы; в частности глоссы могут быть слиты, образуя непарную алаглоссу (alaglossa), либо глоссы и параглоссы слиты вместе, образуя единый непарный придаток – тотаглоссу (totaglossa); алаглоссу или тотаглоссу называют также лигулой (ligula).
У Entognatha нижняя губа неподвижная, так что глоссы и параглоссы слиты, щупики одночлениковые или утрачены [см. Entognatha (1)]. У Triplura глоссы и параглоссы развиты, причем у Microcoryphia параглоссы расщеплены на доли [см. ниже, Microcoryphia (4)]; щупики развиты. У личинок Ephemeroptera глоссы, параглоссы и щупики обычно развиты, но в некоторых таксонах глоссы и/или параглоссы слиты, а их мышцы утрачены (у имаго Ephemeroptera ротовой аппарат утрачен). У стрекоз нижняя губа сильно модифицирована, глоссы и параглоссы утрачены, а щупики модифицированы [см. ниже, Odonata (3c)]. У Idioprothoraca (эмбии и гриллоблаттиды) глоссы, параглоссы и щупики развиты. У большинства Rhipineoptera глоссы, параглоссы и щупики развиты; у уховерток глоссы утрачены и имеются лишь параглоссы [см. ниже, Dermatoptera (3)]. У Zoraptera нижняя губа дистально разделена на пару долей, каждая из которых подразделена на две доли, вероятно соответствующие глоссе и параглоссе; щупики развиты. У всех Acercaria глоссы и параглоссы плохо выражены или отсутствуют; при этом у Parapsocida, Eupsocida, Thysanoptera и части Mallophaga щупики имеются, у прочих Parasita и у Arthroidignatha все парные придатки нижней губы утрачены [см. ниже, Acercaria (3)]. У имаго и личинок насекомых с полным превращением по крайней мере парные глоссы отсутствуют: они либо слиты в непарную алаглоссу, либо совсем утрачены; парные параглоссы бывают развиты или отсутствуют [см. ниже, Metabola (2)]; щупики обычно развиты.
У большинства Hexapoda непарная часть нижней губы расчленена на проксимальную часть – постментум и дистальную часть – прементум. Постментум (postmentum) [называемый также субментум (submentum), или постлабиум (postlabium)] обычно имеет только заднюю (вентральную) стенку и, возможно, является не частью слившихся конечностей, а стернитом. Прементум (praementum) [называемый также ментум (mentum)] представляет собой непарный придаток, подвижно причлененный к переднему краю постментума; глоссы, параглоссы и щупики причленены к прементуму, а идущие от их оснований мышцы крепятся внутри прементума. Подвижное сочленение постментума и прементума хорошо выражено у большинства Amyocerata, но у всех Entognatha оно отсутствует связи с тем, что нижняя губа неподвижно слита с оральными складками (рис. 6.3.1C); так что неясно, является ли наличие поперечного расчленения нижней губы аутапоморфией Hexapoda или лишь Amyocerata.
(3 b) Сравнение. Нижняя губа является уникальным образованием, не встречающимся у других членистоногих. Помимо Hexapoda слияние конечностей IV посторальной пары в непарную нижнюю губу имеется только у симфил [ср.: Symphyla (4) и рис. 5.2.1A]. В отличие от нижней губы Hexapoda, нижняя губа Symphyla сохраняет по всей своей длине продольный медиальный шов, а дистально несет лишь неподвижные выступы, строение которых настолько простое, что неясно, гомологичны ли они глоссам, параглоссам и/или щупикам Hexapoda. У диплопод и пауропод непарное образование, аналогичное нижней губе, образовалось из другой пары конечностей – III посторальной [ср.: Collifera (1)].
(3 c) Число члеников лабиального щупика. По-видимому, исходным для Hexapoda является наличие 3 члеников в лабиальном щупике [в отличие от 5-членикового максиллярного щупика – см. (2c)]. Иногда число члеников лабиального щупика уменьшено благодаря редукции или слиянию тех или иных члеников, иногда щупики утрачены; реже число члеников щупика увеличено благодаря вторичному расчленению некоторых члеников (см. Указатель признаков [1.73]: табл. D). Среди Triplura трехчлениковые щупики имеются у Microcoryphia, а среди Pterygota – у Ephemeroptera, Idioprothoraca, Rhipineoptera, Zoraptera и имаго многих Metabola. У всех Entognatha лабиальные щупики в той или иной мере редуцированы в связи с тем, что вся нижняя губа неподвижно слита с головой; таким образом, неясно, являются ли трехчлениковые лабиальные щупики исходно свойственными Hexapoda или лишь Amyocerata.
У Entognatha лабиальные щупики 1-члениковые или утрачены [см. ниже, Entognatha (1) и Diplura (12)]. У Zygentoma лабиальные щупики выглядят 4-члениковыми [см. ниже, Zygentoma (2)]. У Microcoryphia лабиальные щупики 3-члениковые. У личинок поденок лабиальные щупики обычно 3-члениковые, иногда 2-члениковые [см. ниже, Ephemeroptera (4)]. У стрекоз нижняя губа сильно модифицирована, и ее щупики утратили расчленение [см. ниже, Odonata (3c)]. У Idioprothoraca и Rhipineoptera лабиальные щупики 3-члениковые [см. ниже, Idioprothoraca (6) и Rhipineoptera (3)]. Среди Parametabola 3-члениковые лабиальные щупики сохраняются у Zoraptera. У Acercaria лабиальные щупики подверглись редукции: они имеют не более 2 члеников, и лишь у некоторых трипсов вторично расчленены [см. ниже, Acercaria (3)].
У многих Metabola трехчлениковые лабиальные щупики имеются по крайней мере на стадии имаго. У имаго жуков (Eleuterata) лабиальные щупики обычно 3-члениковые; у личинок жуков 3-члениковые, 2-члениковые или 1-члениковые. У веерокрылых нижняя губа и ее щупики утрачены [см. ниже, Strepsiptera (1), (2), (3)]. У имаго сетчатокрылообразных (Neuropteroidea) лабиальные щупики обычно 3-члениковые, редко 2-члениковые или 4-члениковые; у личинок сетчатокрылообразных лабиальные щупики либо 3-члениковые (рис. 10.5.3A, 10.6.4A и 10.6.6C), либо дополнительно расчленены (рис. 10.4.6A), либо утрачены. У имаго перепончатокрылых число члеников лабиального щупика, вероятно, исходно увеличено до 4; бывает от 1 до 4 члеников; у личинок перепончатокрылых лабиальный щупик 3-члениковый или одночлениковый [см. ниже, Hymenoptera (1)]. У имаго скорпионниц лабиальные щупики 2-члениковые, у блох бывают вторично расчленены или нерасчлененные [см. ниже, Enteracantha (5) и Aphaniptera (1b)]; у личинок скорпионниц и блох лабиальные щупики 2-члениковые или вестигиальные. У имаго двукрылых лабиальные щупики преобразованы в нерасчлененные лабеллумы [см. ниже, Diptera (1B)]; у личинок двукрылых нижняя губа сильно редуцирована и щупики не выражены. У имаго ручейников (Trichoptera) лабиальные щупики исходно 3-члениковые; у некоторых представителей модифицированы, с уменьшенным числом члеников; иногда вестигиальные или утрачены; у личинок лабиальные щупики утрачены. У имаго чешуекрылых (Lepidoptera) лабиальные щупики имеют от 1 до 4 члеников, у большинства 3-члениковые; у личинок лабиальные щупики 2-члениковые.
Исходно лабиальный щупик отличается от максиллярного щупика [см. (2с)], поэтому эволюционной корреляции в их строении не должно быть. Однако в некоторых таксонах насекомых такая корреляция имеется [см. (1b)].
(4) Грудь. Туловище, т.е. отдел, следующий после головы [включающий сегменты начиная с V посторального – см. (11)], разделено на две тагмы – грудь, включающую первые 3 сегмента (V–VII посторальные) и брюшко, включающее остальные 10 сегментов [см. (7) и (8)]. Локомоторные конечности (ноги) сохраняются только на торакальных (т.е. грудных) сегментах; конечности брюшных сегментов редуцированы или модифицированы так, что утратили функцию ног [см. (7)]. В строении трех сегментов груди и их придатков в той или иной мере проявляется собственная эволюционная корреляция [см. (1b)]. В отличие от многих других групп членистоногих, у Hexapoda никогда не меняется число сегментов, входящих в состав груди, т.е. грудь неизменно состоит из трех сегментов (см. Указатель признаков [2.7]). Грудные сегменты отличаются от брюшных не только по строению и наличию конечностей, но и по расположению трахеостий [см. (4b)]. Предложено несколько филогенетических интерпретаций такого строения туловища насекомых [см. (4c)].
(4 a) Состав груди. Три сегмента груди называются соответственно переднегрудь (prothorax), среднегрудь (mesothorax) и заднегрудь (metathorax). Тергит переднегруди называется пронотум (pronotum), тергит среднегруди – мезонотум (mesonotum), тергит заднегруди – метанотум (metanotum); стерниты называются соответственно простернум (prosternum), мезостернум (mesosternum) и метастернум (metasternum), а плейральные области каждого из этих сегментов – соответственно проплевры (propleura), мезоплевры (mesopleura) и метаплевры (metapleura). Каждый из трех сегментов груди несет пару локомоторных конечностей, называемых соответственно передние ноги, средние ноги, и задние ноги [см. (5)].
(4 b) Расположение трахеостий. Трахеостии [парные латеральные – см. (9)] по-разному расположены на груди и брюшке. В примитивном случае имеются следующие пары трахеостий (рис. 6.1.1 и 6.3.1A). Первая пара, стеноторакальная – между переднегрудью и среднегрудью (сохраняется у Diplura и Amyocerata); вторая пара, мезоторакальная – на среднегруди (сохраняется у Diplura и Protura); третья пара пара, криптоторакальная – между среднегрудью и заднегрудью (сохраняется у Diplura и Amyocerata); четвертая пара, метаторакальная – на заднегруди (сохраняется у Diplura и Protura) и V–XII пары, абдоминальные – на каждом из I–VIII сегментов брюшка (все или некоторые из них сохраняются у Diplura и Amyocerata) [о происхождении терминов «стеноторакальная» и «криптоторакальная» – см. (4с)]. Таким образом, могут присутствовать 2 пары межсегментарных трахеостий груди (I и III пары), 2 пары сегментарных трахеостий груди (II и IV пары) и 8 пар сегментарных трахеостий брюшка (V–XII пары); все или часть из них могут служить дыхальцами. У Amyocerata имеется максимум 2 пары межсегментарных дыхалец на груди и 8 пар сегментарных дыхалец на брюшке, иногда часть из них утрачена [см. ниже, Amyocerata (2)]. Среди Entognatha, у двухвосток имеется до 4 пар дыхалец на груди и до 7 пар дыхалец на брюшке [см. ниже, Diplura (8)], иногда часть из них утрачена; у бессяжковых могут присутствовать 2 пары сегментарных дыхалец на груди (не гомологичных межсегментарным грудным дыхальцам Amyocerata), иногда они утрачены [см. ниже, Protura (9)]; у ногохвосток первичные трахеостии утрачены [см. ниже, Collembola (4)]. Следует учесть, что иногда дыхальца бывают смещены на соседний сегмент, что затрудняет их гомологизацию [см. (9)].
(4 c) Гипотезы о происхождение груди. Помимо Hexapoda, различие между V–VII посторальными сегментами (соответствующим груди) и остальными туловищными сегментами (соответствующими брюшку) в той или иной форме выражено у Collifera: 3 сегмента, соответствующие груди, несут 3 первые пары ног; остальные туловищные сегменты у личинок диплопод и пауропод не имеют ног [ср.: Collifera (3) и рис. 5.3C, 5.4.1C], а у взрослых диплопод несут по две пары ног [ср.: Diplopoda (3) и рис. рис. 5.3A]. У симфил таких четких различий между этими отделами тела нет, но первые три сегмента туловища реже, чем другие, имеют по два тергита на одну пару ног [ср.: Symphyla (5) и рис. 5.2.3]. Некоторые авторы рассматривают это как синапоморфию Hexapoda и Progoneata (см. Atelocerata: «Классификации» III), связывая это с гипотезой о происхождении Hexapoda от Myriapoda. Однако если исходить из того, что Myriapoda – голофилетический таксон, то приходится делать вывод о независимой дифференциации первых трех туловищных сегментов у Progoneata и у Hexapoda.
Было высказано предположение, что в состав груди Hexapoda вошли не 3, а 6 исходных сегментов, объединенных в три диплосегмента (подобно тому, как это имеет место у двупарноногих многоножек); из этих шести сегментов I, III и V сегменты подверглись редукции и утратили ноги, а II, IV и VI сегменты остались развитыми. Для каждого из гипотетических редуцированных сегментов были предложены собственные названия (Verhoeff 1903): микроторакс (microthorax) – шейный сегмент между головой и переднегрудью; стеноторакс (stenothorax) – сегмент между переднегрудью и среднегрудью; криптоторакс (cryptothorax) – сегмент между среднегрудью и заднегрудью. При этом предполагалось, что каждый сегмент исходно имел одну пару трахеостий, так что межсегментарные трахеостии (т.е. трахеостии I и III пар) относятся к вестигиям стеноторакса и криптоторакса соответственно [см. (4b)]. Автор этой гипотезы считал, что тергиты и стерниты стеноторакса и криптоторакса, подвижно сочлененные с соседними тергитами и стернитами, сохранились у Dicellurata и Embioptera. Однако обособленные склериты, который он считал стернитами стеноторакса и криптоторакса у эмбий (Verhoeff 1904a), являются спинастернумами и несут первую и вторую спины (рис. 8.2.2A), тогда как обособленные склериты, который он считал стернитами стеноторакса и криптоторакса у двухвосток (Verhoeff 1904c), расположены позади оснований первой и второй спин (рис. 6.3.1C, 6.3.2D); если верно предположение о гомологии спин у Diplura и у Pterygota [см. Pterygota (3)], то склериты, принимавшиеся за стерниты стеноторакса и криптоторакса у Dicellurata и Embioptera, негомологичны. Представление о том, что шейные склериты произошли из особого сегмента микроторакса, связано с тем, что у Dicellurata, помимо Y-образных гребней на переднегруди, среднегруди и заднегруди, имеется еще один Y-образный гребень между головой и переднегрудью, где он не несет спину и не выполняет никаких функций, так что выглядит как вестигиальный Y-образный гребень, принадлежащий вестигию микроторакса (рис. 6.3.1C).
Некоторые авторы считали, что ноги промежуточных сегментов груди не исчезли, а вошли в состав существующих трех пар ног. Тазик ноги [см. (5a)] бывает разделен продольным швом на переднюю и заднюю части; было высказано предположение, что задняя часть тазика исходно принадлежала ноге следующего, промежуточного, сегмента груди; по недоразумению эта задняя часть тазика была названа «мерон» («meron»), поскольку у крылатых насекомых она расположена под эпимероном [см. ниже, Pterygota (3)] (Walton 1900); однако слово «meron» (древнегреческое «μηρος») означает «бедро», т.е. является переводом термина «femur», который относится к другой части ноги [см. (5a)]. В последствие предположение о срастании ног разных пар было отвергнуто, а термин «мерон» стали применять к задней части тазика, отделенной продольным швом и вмещающей места прикрепления тех плейро-коксальных и/или терго-коксальных мышц, которые принимают участие в движении крыла и в связи с этим увеличены [см. ниже, Pterygota (4Abγ)]. На самом деле у тазиков разных насекомых наличие и положение продольного шва (который может служить механической опорой для плейро-коксального мыщелка) по-разному сочетается с местами прикрепления мышц, так что термин «мерон» не относится к какой-либо конкретной части тазика насекомых.
Гипотеза о происхождении сегментов груди из диплосегментов связана с предположением, что Hexapoda произошли от Myriapoda, в то время как наличие диплосегментов является исходным свойством многоножек [ср.: Myriapoda (3)].
Возможно и другое объяснение того, что в груди Hexapoda три нормально развитых сегмента чередуются с элементами еще трех сегментов. У членистоногих имеются регуляторные гены т.н. «парного правила», которые в эмбриогенезе экспрессируются в зонах, каждая из которых соответствует двум будущим сегментам тела; вероятно, наличие такого механизма, регулирующего онтогенез, может приводить к тому, что в ходе эволюции происходит частичное или полное удвоение сегментов (Minelli & Bortoletto 1988).
(5) Особенности ног. Ноги [в количестве 3 пар, только на груди – см. (4)] имеют особое строение телоподита [см. Euarthropoda (3a)]: помимо дистальных отделов – лапки и претарсуса [см. Atelocerata (3)] телоподит включает лишь вертлуг, бедро и цельную голень [см. (5a)]. Несмотря на то, что те же названия частей ноги используются и для других групп членистоногих, данное строение свойственно только Hexapoda [см. (5b)].
(5 a) Строение ноги (рис. 6.1.2). В строении ног всех трех пар проявляется эволюционная корреляция, обычно хорошо выраженная [как и у ног многих других членистоногих – см. Gnathopoda (4)].
Коксоподит [см. Euarthropoda (3a)], называемый здесь тазик, или кокса (coxa), на всех грудных ногах лишен гнатобазы (как и на большинстве туловищных ног других членистоногих). Положение тазика варьирует от латерального до вентрального (некоторые авторы необоснованно считают латеральное или вентральное положение тазиков признаком, объединяющим насекомых с той или иной группой членистоногих).
Обычно (но не всегда) тазик сочленяется с туловищем двумя мыщелками – дорсо-латеральным и вентро-медиальным (при разной форме туловища ось вращения тазика может быть не перпендикулярной, а наклонной по отношению к оси тела; здесь слова «дорсальный» и «вентральный» употребляются так, как если ось вращения тазика перпендикулярна оси тела).
Дорсо-латеральный мыщелок соединяет тазик с плейритом [точнее, с катаплейритом – см. (5b)] и, вероятно, гомологичен у всех Hexapoda.
Вентро-медиальный мыщелок соединяет тазик либо со стернитом, либо с трохантином. Трохантин (trochantinus), если имеется, представляет собой более или менее обособленный участок катаплейрита, прилегающий к переднему краю основания тазика. Соединение тазика со стернитом бывает либо непосредственным, либо посредством обособленного склерита. Иногда тазик сочленен и с трохантином, и со стернитом, и, таким образом, имеет три мыщелка – один дорсо-латеральный и два вентро-медиальных; в этом случае его подвижность обеспечивается либо подвижностью трохантина, либо подвижностью сочленовного стернального склерита.
У Diplura и Protura трохантины отсутствуют, и тазики сочленяются со стернитами (рис. 6.3.1C). У Collembola вентральная сторона тела не склеротизована, так что вентро-медиальные мыщелки тазиков не выражены (рис. 6.4.1E). У Triplura тазики сочленяются с трохантинами (рис. 6.7.1A и 6.8.1A). У Ephemeroptera (у личинок, субимаго и имаго) на переднегруди трохантины имеются или отсутствуют, а на среднегруди и заднегруди трохантины всегда отсутствуют, и тазики сочленяются со стернитами либо непосредственно, либо посредством особых подвижных склеритов (рис. 7.2.3:I). У Odonata трохантины отсутствуют, но у оснований средних и задних ног личинки могут быть маленькие склериты, предположительно гомологичные трохантинам (Matsuda 1970). У большинства Neoptera трохантины развиты; у некоторых из них развито также сочленение тазика со стернитом. У цикадовых оба сочленения развиты на задних ногах [см. ниже, Euhomoptera (3) и рис. 9.7.3D)]. У имаго многих Metabola средний и задний тазики сочленяются и с трохантином, и со стернитом, иногда только со стернитом; при этом иногда стернит в местах сочленений с тазиками вытянут, образуя пару сочленовных выростов [см. Eumetabola (3) и рис. 10.8.1A]; у блох эти выросты обособлены в виде пары подвижных сочленовных склеритов [см. ниже, Aphaniptera (1a)].
К тазику посредством подвижного двухмыщелкового сочленения причленен сравнительно короткий вертлуг, или трохантер (trochanter), направленный латерально и двигающийся дорсовентрально относительно тазика. К вертлугу малоподвижно причленено относительно длинное бедро, или фемур (femur), обычно имеющее то же направление, что и вертлуг. К бедру подвижно причленена голень, или тибиа (tibia) [состоящая из слитых пателлы и телотибии – см. (5b)]; бедро и голень образуют единственное коленное сочленение, которое обычно является двухмыщелковым (см. ниже, Amyocerata: «Классификации» I) и позволяет голени совершать дорсовентральные движения в пределах от состояния вытянутого в одну линию с бедром до согнутого вентрально и прижатого к бедру (аналогично коленному суставу позвоночных). К голени причленена лапка, или тарсус (tarsus) [см. Atelocerata (3)]; у Entognatha лапка нерасчлененная, у Amyocerata она имеет до 5 члеников [см. ниже, Entognatha (3) и Amyocerata (5)]. У многих насекомых лапка преимущественно направлена в одну линию с голенью (рис. 7.2.1); у некоторых крылатых насекомых лапка преимущественно направлена перпендикулярно голени и служит ступней. К лапке причленен претарсус (praetarsus), сгибающийся вентрально [см. Atelocerata (3)] и несущий коготки [см. (6)].
Тазик (кокса), будучи коксоподитом, двигается мышцами, крепящимися в туловище; различные коксальные мышцы могут крепится к тергиту, плейриту, стерниту, мезодермальным сухожилиям и/или кутикулярным аподемам – к фурке, спине и/или плейральной аподеме [см. ниже, Pterygota (3)]. Вертлуг (трохантер), будучи первым члеником телоподита, двигается мышцами, крепящимися в тазике и в туловище; различные трохантеральные мышцы могут крепится к тергиту, плейриту, плейральной аподеме и/или фурке. В отличие от Myriapoda (Manton 1955, 1958, 1961, 1965, 1966), у Hexapoda туловищные коксальные и трохантеральные мышцы каждой ноги крепятся преимущественно в пределах своей половины сегмента: за редким исключением, они не распространяются по другую сторону от медиальной плоскости и не тянутся в другие сегменты (Matsuda 1970). Прочие членики ноги двигаются только мышцами, крепящимися в предыдущих члениках: к основанию бедра идут мышцы только из вертлуга; к основанию голени могут идти мышцы из вертлуга и бедра, к основанию лапки – из голени, к единственной аподеме претарсуса – из бедра и голени (рис. 6.1.2).
Других члеников ноги нет. Бывает лишь явно вторичное расчленение или слияние названных члеников; например у Collembola и у личинок некоторых Pterygota голень и лапка сливаются в один членик – тибиотарсус (см. Указатель признаков [2.49]).
Если нога имеет предпочтительное место отлома, то этим местом обычно является малоподвижное сочленение вертлуга с бедром: при опасности у некоторых насекомых в этом месте может происходить автоспазия (когда нога легко отламывается) или автотомия (когда нога самопроизвольно отбрасывается) (Maruzzo et al. 2005).
(5 b) Гомология частей ноги. Существуют разногласия по поводу того, что считать истинным первым члеником ноги тех или иных членистоногих. Согласно субкоксальной теории происхождения плейритов, плейрит грудного сегмента насекомых возник благодаря тому, что в состав стенки туловища вошел либо отделившийся от тазика проксимальный участок, либо один или несколько члеников, исходно имевшихся проксимальнее тазика: предельно укоротившиеся членики образовали вокруг основания ноги несколько концентрических склеротизованных колец или полуколец, разделенных концентрическими швами. У разных насекомых можно усмотреть разное число таких склеротизованных полуколец (рис. 6.7.1A), но иногда весь плейрит представлен цельным склеритом. Принято считать, что относительно постоянным для насекомых является паракоксальный шов, разделяющий плейрит на наружное полукольцо – анаплейрит, или анаплейрон (anapleuron) и внутреннее полукольцо – катаплейрит, или катаплейрон (katapleuron) [о более дробном делении плейрита у крылатых насекомых – см. ниже, Pterygota (3)]. По мнению некоторых авторов у Hexapoda бывает развита субкокса – членик проксимальнее тазика; в частности, указывали на наличие субкокс у ногохвосток; однако детальное сравнительное исследование мускулатуры, кутикулы и мезодермального эндоскелета показало, что в данном случае за субкоксу передних ног принимали несклеротизованную плейральную выпуклость, а за субкоксы средних и задних ног – обособленные проксимальные части тазиков (Manton 1972) (рис. 6.4.1A).
Вероятно, голень Hexapoda образована в результате слияния двух исходных члеников – пателлы (patella) и телотибии (telotibia); иногда телотибию называют голенью, а ее вместе с пателлой – метатибией (metatibia). У многих поденок (Ephemeroptera) и стрекоз (Odonata) граница между пателлой и телотибией представляет собой пателло-тибиальный шов, идущий наискось от наружно-проксимальной части голени ковнутри-дистально (рис. 7.2.1, 7.4.1A, E); этот шов может полностью опоясывать голень, либо прерван на задней стороне голени; у личинок поденок и стрекоз он обычно имеет вид тонкой борозды, а у имаго поденок и стрекоз представляет собой лишь неясное пологое вдавление; у поденок пателло-тибиальный шов исходно имеется только на средних и задних ногах [см. ниже, Ephemeroptera (8)], у стрекоз на всех ногах; у части поденок и стрекоз пателло-тибиальный шов утрачен; во всех случаях пателла и телотибиа соединены совершенно неподвижно. У других Hexapoda (Entognatha, Triplura и Neoptera) пателло-тибиальный шов почти не выражен или совсем утрачен.
В отличие о Hexapoda, у большинства других членистоногих, в том числе у всех многоножек, имеется подвижная пателла, обеспечивающая сгибание ноги не по одному коленному сочленению, а по двум – феморо-пателлярному и пателло-тибиальному (рис. 4.8). Помимо этого, каждая из групп многоножек имеет собственный план строения ноги [ср.: Symphyla (11), Diplopoda (5), Pauropoda (6) и Chilopoda (7)]: у Chilopoda имеется вертлуг, вероятно, гомологичный вертлугу Hexapoda; у Symphyla и Pauropoda вместо вертлуга и бедра имеется единый членик – трохантерофемур (обычно называемый трохантером или фемуром); у Diplopoda вместо вертлуга имеется два членика – трохантер и префемур (иногда называемые иначе).
(5 c) Модификации ног. У большинства насекомых все три пары ног ходильные или бегательные. В отдельных таксонах ноги всех или отдельных пар видоизменены и приобрели новую функцию – прыгательную, плавательную, копательную, фильтрующую, хватательную или иную; при этом исходная ходильная специализация либо сохраняется, либо утрачивается.
При хватательной специализации передней ноги бедро и голень используются как зажим, удерживающий добычу; тазик может быть либо не измененным и коротким, либо удлинен так, что позволяет выдвигать бедро вперед. Такая модификация передних ног имеется у личинок некоторых поденок (Ephemeroptera), у богомолов [см. ниже, Raptoriae (1)], у некоторых тараканов (Neoblattariae) и некоторых прыгающих прямокрылых (Saltatoria), у Mantophasma/f=Raptophasma/g (см.ниже, Spectra: «Классификация»), у некоторых клопов (Heteroptera), у некоторых двухоботных сетчатокрылых (см. Birostrata: «Классификации»: Dictyneuroptera и Berotha/fg) и у некоторых других хищных насекомых. При этом лапка и коготки могут сохранять исходное строение, так что передняя нога совмещает хватательную функцию с исходной ходильной (у богомолов и многих других). У некоторых хищных клопов (в составе Hydrocorisae, Reduvius/fg и др.) и у мантиспид (Dictyneuroptera) тарсус и претарсус передней ноги специализированы как часть хватательного аппарата или редуцированы, так что передняя нога утратила локомоторную функцию, подобно ногочелюстям хилопод и ракообразных.
В некоторых случаях ноги определенных пар или все ноги имеют упрощенное строение, при котором некоторые членики утрачены или слиты друг с другом [см. (5a)]; в некоторых случаях ноги утрачены (см. Указатель признаков [2.38]–[2.98]).
(6) Коготки претарсуса. Претарсус [самый дистальный отдел ноги, сгибаемый унгуитрактором – см. (5) и Atelocerata (3)] имеет два боковых коготка (ungues) – т.е. два коготка, причлененных по бокам исходного срединного коготка (дактилоподита); таким образом, претарсус исходно несет 3 коготка (рис. 6.1.2). Среди Entognatha дактилоподит и оба боковых коготка имеются у Diplura, тогда как у ногохвосток и протур боковые коготки утрачены [см. ниже, Ellipura (2)]. У Diplura оба боковых коготка причленены к основанию дактилоподита и двигаются вместе с ним (рис. 6.3.1D). Среди Amyocerata дактилоподит и оба боковых коготка имеются у большинства Zygentoma (рис. 6.7.3A), тогда как у Microcoryphia и Pterygota дактилоподит редуцирован и обычно имеются лишь два боковых коготка [см. ниже, Microcoryphia (8) и Pterygota (5)]. У Amyocerata все коготки (три коготка у Zygentoma или два коготка у Microcoryphia и Pterygota) вентрально сочленены с унгуитрактором [см. Atelocerata (3)], а дорсально подвижно причленены к унгуиферу (unguifer), который представляет собой неподвижный выступ на вершине последнего членика лапки (рис. 6.7.3A, 7.2.4D). В строении двух боковых коготков одной ноги проявляется собственная эволюционная корреляция [кроме того, имеется эволюционная корреляция в строении коготков трех пар ног – см. (5)]. Иногда один из боковых коготков редуцируется или происходят иные преобразования, в результате чего имеется лишь один коготок; иногда коготки утрачены (см. Указатель признаков [2.58]).
На основании того, что у Ellipura боковые коготки отсутствуют, некоторые авторы не считают наличие боковых коготков исходным признаком Hexapoda.
Среди Atelocerata, помимо Hexapoda, два боковых коготка имеются также у Pauropoda; один боковой коготок имеется у Symphyla.
У некоторых других наземных членистоногих (онихофоры и некоторые паукообразные) нога также заканчивается двумя коготками; в отличие от Atelocerata, у этих животных коготки способны к активному мышечному разгибанию [см. Gnathopoda (4), Onychophora (1), Chelicerata: «Классификации» III: Arachnida (3)].
(7) I–IX сегменты брюшка и их придатки. Из десяти сегментов брюшка (уромеров) первые 9 сегментов исходно имеют особое строение, в котором проявляется эволюционная корреляция. Последний, десятый, уромер исходно иное особое строение, не имеющее ничего общего со строением предыдущих сегментов [см. (8)].
В примитивном случае стернит каждого из первых девяти уромеров несет на заднем крае пару подвижных стерностилей, а между ними пару кинетапофизов (рис. 6.6.3).
Стерностиль (sternostylus) представляет собой подвижный одночлениковый придаток палочковидной формы. К основанию стерностиля подходит мышца, неподвижный конец которой крепится к стерниту или к стилигеру (см. ниже). Внутри самого стерностиля мышц никогда не бывает. Стерностиль может нести на вершине неподвижный склеротизованный заостренный вырост, далее называемый апикальный шип. Иногда апикальный шип называют тарселлусом (tersellus), что связано с предположением о его соответствии дистальному отделу конечности – тарсусу. Однако, в отличие от отдела конечности, апикальный шип представляет собой неподвижный кутикулярный вырост, не имеющий полости внутри.
Обычно стерностиль называют просто «стилусом», или «грифельком». В некоторых случаях это приводит к недоразумениям, потому что термин «стилус» (stylus), помимо стерностиля, применяют и к другим полочковидным придаткам самого разного расположения и происхождения, таким как отчлененные придатки девятого тергита брюшка у некоторых жуков и др. (см. Указатель русских терминов и Указатель латинских терминов).
Кинетапофиз (kinetapophysis) представляет собой либо мембранозный втяжной пузырек (vesicula retractilis), способный вворачиваться и втягиваться в тело, либо невтягивающийся подвижный отросток. К основанию кинетапофиза, как и к основанию стерностиля, подходит мышца, неподвижный конец которой крепится к стерниту или к стилигеру.
Стернит может быть цельным, либо разделен поперечным стилигеральным швом на переднюю часть – престилигер (praestyliger) и заднюю часть – стилигер (styliger). В этом случае все мышцы, идущие к стерностилям и кинетапофизам крепятся в пределах стилигера. Стилигер может сильно выступать назад в виде пластины или пары пластин, на заднем крае которых находятся стерностили и кинетапофизы. Единый непарный стилигер, несущий левые и правые стерностили и/или кинетапофизы, можно называть бистилигером (bistyliger), а обособленные левый или правый стилигеры, каждый из которых несет один стерностиль и/или один кинетапофиз, можно называть унистилигером (unistyliger).
На прегенитальных сегментах брюшка (т.е. на I–VII уромерах) стерностили и кинетапофизы развиты только у первичнобескрылых насекомых – Triplura и Diplura. Кинетапофизы этих сегментов обычно представляют собой втяжные пузырьки, лишь у части Diplura кинетапофизы I уромера являются невтягивающимися придатками [см. ниже, Rhabdura (1) и Prodicellura (2)]. У Triplura-Microcoryphia на некоторых уромерах вместо одной пары втяжных пузырьков имеется две пары втяжных пузырьков [см. ниже, Microcoryphia (9)]. Стилигеральные швы выражены только у части Triplura (у всех Microcoryphia и некоторых Zygentoma), тогда как у всех Diplura и многих Zygentoma стерниты брюшка цельные. У многих Diplura и Triplura стерностили и/или кинетапофизы тех или иных прегенитальных сегментов утрачены [см. ниже, Diplura (5) и Triplura (6)]. У Pterygota стерностили и кинетапофизы всех прегенитальных сегментов утрачены [см. ниже, Pterygota (6)]; у гриллоблаттид на первом стерните блюшка имеется непарный втяжной пузырек, вероятно не гомологичный парным кинетапофизам [см. ниже, Notoptera (2)].
На генитальных сегментах брюшка (т.е. на VIII–IX уромерах) у самки кинетапофизы либо представляют собой створки яйцеклада, либо утрачены [см. (13) и Amyocerata (7)]. У Triplura стерностили генитальных сегментов сходны со стерностилями прегенитальных сегментов; у крылатых насекомых стерностили сохраняются только на IX уромере [см. ниже, Pterygota (6)]. У самцов поденок стерностили IX уромера сохраняют подвижность и преобразованы во вторичночленистые половые придатки [см. ниже, Ephemeroptera (15a)]; у всех прочих крылатых насекомых утрачены мышцы, подходящие к стерностилям [см. ниже, Metapterygota (5)]. У многих крылатых насекомых утрачены все производные стерностилей и кинетапофизов.
В ходе онтогенеза стерностили и кинетапофизы тех или иных сегментов брюшка либо закладываются в эмбриогенезе, либо появляются лишь в постэмбриогенезе при тех или иных линьках.
Предполагают, что унистилигеры, стерностили и кинетапофизы являются вестигиями утраченных абдоминальных ног, но их гомология с частями ноги неясна. Стилигеры часто называют «кокситами», что связано с предположением об их гомологии с коксам ног. Однако, в отличие от кокс, стилигеры являются не обособленными члениками, а составляют часть вентральной стенки тела; они всегда расположены не по бокам, а позади стернита; стилигеры неподвижны, и мышцы из туловища к ним не подходят. Для некоторых ископаемых насекомых – Triassomachilis uralensis Sharov 1948, Permuralia maculata (Kukalova-Peck et Sinitshenkova 1992 [Uralia]) и некоторых других – были ошибочно описаны членистые стерностили; в действительности T. uralensis представляет собой личинку поденки и не имеет стерностилей, а у P. maculata имеются нечленистые отростки, похожие на стерностили. Стерностили исходно нечленистые; лишь их производные – гоностили поденок – вторично разделены на безмышечные членики [см. ниже, Ephemeroptera (15a)].
Парные стилусы и втяжные пузырьки, внешне похожие на стерностили и кинетапофизы Hexapoda, имеются у симфил, где они располагаются между основаниями ног [ср.: Symphyla (6) и рис. 5.2.1E]; в отличие от Hexapoda, у Symphyla к стилусам мышцы не подходят, а к пузырькам подходят мышцы, начинающиеся не от стернита или стилигера, а от коксальных аподем. Втяжные пузырьки, кроме того, имеются у некоторых других многоножек [ср.: Progoneata (5)] – на тазиках ног или между ногами у некоторых диплопод [ср.: Diplopoda (8) и рис. 5.3F–G] и на утратившем конечности коллюме у пауропод [ср.: Pauropoda (4) и рис. 5.4.1D]. Помимо Atelocerata, втяжные пузырьки имеются у некоторых почвенных хелицеровых на вестигиях конечностей опистосомы или на утратившей конечности опистосоме и в основаниях ног некоторых онихофор (см. Указатель признаков [3.36] и рис. 4.2D). Из этого можно заключить, что сходные по строению втяжные пузырьки несколько раз независимо возникали на вентральной стороне тела у разных членистоногих, перешедших к почвенному или напочвенному образу жизни.
На основании того, что строение придатков брюшка имеет явное сходство у Diplura (относящихся к Entognatha) и у Triplura (относящихся к Amyocerata), можно заключить, что это строение является исходным для Hexapoda. Эти же черты строения прослеживаются и у Pterygota, так что не возникает сомнений в том, что они исходно свойственны всем Amyocerata. Однако дискуссии вызвал вопрос о происхождении конечностей брюшка Ellipura (см. «Классификации» III).
(8) Десятый сегмент брюшка. Брюшко исходно состоит из 10 сегментов (уромеров); то есть все тело имеет 17 посторальных сегментов, включая 4 головных, 3 грудных и 10 брюшных [см. (4) и (11)]. Десятый уромер, на котором располагается анальное отверстие, не имеет структур, присущих первым девяти уромерам [см. (7)]. Среди Entognatha десятый уромер имеется только у Diplura, тогда как у ногохвосток и протур он полностью утрачен [см. ниже, Ellipura (3)]. Среди Amyocerata десятый уромер имеется у всех представителей, но у некоторых он упрощен и/или уменьшен. Десятый уромер может нести парные подвижные придатки, называемые «церками», но имеющие разное строение у Diplura и у Amyocerata [см. (8a)]. По поводу строения и гомологии вершины брюшка высказывались различные мнения [см. (8b)].
(8 a) Каудалии и псевдокаудалии. У многих Amyocerata и у всех Diplura десятый уромер несет пару придатков, подвижно причлененных к его заднему краю. Эти придатки принято называть «церками», считая их гомологичными. Однако строение этих придатков принципиально различается у Amyocerata и у Diplura. У Amyocerata придатки десятого уромера представляют собой каудалии; их исходно не два, а три (пара церков и непарный парацерк), и они исходно занимают дорсальное положение, так что непарная каудалия расположена дорсальнее анального отверстия; все мышцы, идущие к основаниям каудалий, включая аддукторы церков, крепятся к тергиту [см. ниже, Amyocerata (8) и рис. 7.2.7]; в большинстве случаев каудалии представлены только парными церками [см. ниже, Metapterygota (4)]. У Diplura придатки десятого уромера представляют собой псевдокаудалии; псевдокаудалий всегда только две, и, в отличие от каудалий, они связаны с вентральной стороной тела, поскольку их мышцы-аддукторы крепятся вентральнее продольных стернальных мышц [см. ниже, Diplura (9) и рис. 6.3.2A–B].
Помимо каудалий и псевдокаудалий, на конце брюшка насекомых и других членистоногих бывают развиты сходные с ними парные торчащие назад придатки другого строения и независимого происхождения.
У Chilopoda последняя пара ног направлена назад и при ходьбе волочится; у многих Chilopoda-Epimorpha ноги этой пары сильно отличаются от ходильных ног, не имеют претарсуса и торчат назад прямо, так что напоминают церки [ср.: Chilopoda (7)]. По аналогии с этим существует широко распространенное мнение, что церки всех насекомых (и Amyocerata, и Diplura) произошли из конечностей последнего сегмента тела [см. (8b)]. Было даже высказано предположение о непосредственной гомологии между церкоподобными задними ногами хилопод и церками насекомых, что связано с представлением о родстве Hexapoda с Chilopoda и о том, что среди Chilopoda наиболее примитивными являются Epimorpha (Verhoeff 1904c). Однако, согласно современным представлениям о филогении хилопод, Epimorpha далеки от общего предка Chilopoda, а Chilopoda относятся к голофилетическому таксону Myriapoda и поэтому не могут иметь непосредственного родства с Hexapoda [ср.: Myriapoda (1) и Chilopoda: «Классификации» III]. Церкоподобные ноги хилопод, как и другие ноги, состоят из настоящих члеников с мышцами, в отличие от каудалий и псевдокаудалий, которые либо не расчленены, либо имеют лишь вторичное расчленение без внутренних мышц.
У Symphyla последний тергит туловища несет направленные назад прядильные придатки, которые принято называть «церками»; некоторые авторы считают их гомологичными церкам насекомых [ср.: Symphyla (8)]. В отличие от церкоподобных ног хилопод, прядильные придатки симфил не могут быть сериальными гомологами ног, поскольку они находятся на тех же сегментах, что и развитые ходильные ноги. Доводом в пользу гомологии прядильных придатков симфил и церков насекомых считают наличие протоков шелкоотделительных желез в хвостовых придатках Diplura-Prodicellura, которые считают гомологичными церкам [см. ниже, Prodicellura (3)]. Однако хвостовые придатки Diplura (псевдокаудалии) двигаются вентральными мышцами, тогда как прядильные придатки симфил двигаются мышцами, идущими от тергита. В отличие от псевдокаудалий Diplura и церков Amyocerata, прядильные придатки симфил находятся не на конце тела, а на его дорсальной стороне, и позади них имеется еще один отдел туловища – тельсон. Представление о гомологии псевдокаудалий Prodicellura и прядильных придатков Symphyla связано с гипотезой о родстве Hexapoda и Symphyla (которая противоречит представлению о голофилии Myriapoda) и с гипотезой о происхождении всех Hexapoda из Diplura (которая противоречит представлению о голофилии Entognatha).
Альтернативное предположение состоит в том, что церки Hexapoda гомологичны фурке ракообразных [ср.: Eucrustacea (5)]. Фурка ракообразных не может быть гомологичной ни церкоподобным ногам хилопод, ни прядильным придаткам симфил, поскольку находится на тельсоне, т.е. позади зоны добавления новых сегментов.
Некоторые авторы принимали за церки удлиненные подвижные парапрокты личинок стрекоз [см. ниже, Odonata (10)], урогомфы на девятом уромере личинок жуков [см. ниже, Elytrophora (12)] и другие придатки, находящиеся на конце тела и направленные назад.
Вопрос о том, какие именно из придатков членистоногих следует называть «церками», нужно решать исходя из первоначального употребления этого термина. Снодграсс (Snodgrass 1909: 572) указывает авторство этого термина как Kirby & Spense 1826. Эти авторы дали определения различным морфологическим терминам, в том числе названиям анальных придатков («appendices»), среди которых фигурируют «cerci» (примеры – Blatta и Gryllus), «caudulae» (примеры – Lepisma, Machilis, Ephemera) и «forceps» (пример – Forficula) (Kirby & Spense 1826, vol. 3: 391–393). Таким образом, термин «церки» («cerci», в единственном числе «cercus») исходно применяется к парным каудалиям Amyocerata.
(8 b) Дискуссии о числе сегментов брюшка. Существует широко распространенное мнение, что Hexapoda произошли от каких-то членистоногих (Myriapoda, Eucrustacea или др.), имевших больше 17 посторальных сегментов, и что наиболее примитивные представители Hexapoda сохраняют остатки XI или даже XII сегмента брюшка.
Однако в действительности ни у каких Hexapoda нет ни мышечных комплексов, ни конечностей, которые можно было бы интерпретировать как принадлежащие утраченным сегментам, располагавшимся позади десятого сегмента брюшка. Нет также и следов их нервных ганглиев [см. Metameria (2)]; более того, нервный ганглий, иннервирующий десятый уромер, всегда слит с ганглиями IX и VIII уромеров в единый синганглий, обычно расположенный в области VIII уромера; так что в брюшке Hexapoda имеется не более 8 ганглиев вентральной нервной цепочки; иногда число ганглиев еще меньше из-за слияния их друг с другом.
Авторы, открывшие протур (Silvestri 1907, Berlese 1909), считали их промежуточной формой между Myriapoda и Hexapoda, сохраняющей 12 сегментов брюшка. Однако более правдоподобное объяснение строения протур состоит в том, что у них, наоборот, утрачен десятый сегмент, а иллюзия многосегментности создается за счет наличия трех вторичных колец, отделившихся от восьмого сегмента брюшка (Lameere 1935, Hennig 1969, 1981) [см. ниже, Protura (6)].
Существует широко распространенная теория о том, что у Amyocerata трикаудальный склерит (включающий эпипрокт) и парапрокты являются остатками одиннадцатого сегмента брюшка, а церки являются его видоизмененными конечностями (Snodgrass 1931, 1935). Против этой интерпретации свидетельствуют следующие факты. Отдел, считающийся одиннадцатым сегментом брюшка, не имеет собственной мускулатуры, в отличие от настоящих сегментов брюшка, каждый из которых имеет собственный набор продольных и дорсовентральных мышц или их производных [см. Euarthropoda (1)]. Мышцы, идущие к церкам, у всех Amyocerata крепятся к тергиту десятого сегмента брюшка, и нет ни одной мышцы, которая соединяла бы церки с парапроктами или эпипроктом – то есть с тем, что считают одиннадцатым сегментом брюшка. Если считать парапрокты и церки относящимися к одиннадцатому сегменту брюшка, то получается, что десятый сегмент брюшка тоже не имеет собственного набора мышц, поскольку все мышцы от тергита десятого сегмента идут к склеритам, считающимся частями одиннадцатого сегмента. Представление о церках Amyocerata как о производных пары конечностей противоречит единству строения и наличию эволюционной корреляции у всех трех каудалий – церков и парацерка [см. ниже, Amyocerata (8)].
Представление о существовании одиннадцатого сегмента брюшка возникло в связи с тем, что наиболее примитивным таксоном среди Amyocerata считались Microcoryphia, которые в действительности имеют весьма модифицированное строение брюшка [см. ниже, Microcoryphia (10)]: десятый тергит брюшка замкнут в кольцо (так что кажется целым сегментом), а каудалии и парапрокты неподвижно слиты друг с другом позади него, так что их цельное основание кажется еще одним, одиннадцатым сегментом (рис. 6.8.2). Помимо Microcoryphia, десятый тергит брюшка сомкнулся вентрально и оттеснил парапрокты назад у некоторых других насекомых [см. ниже, Amyocerata (8b)]. Однако у многих насекомых (у Zygentoma, Ephemeroptera и др.) десятый тергит брюшка не столь сильно заходит на вентральную сторону, так что парапрокты причленяются не к нему, а к девятому стерниту и находятся вентральнее десятого тергита (рис. 6.7.2A, C, E, 7.2.7, 8.3.2A); в этом случае принадлежность парапроктов десятому сегменту очевидна.
Помимо Microcoryphia, нечто внешне похожее на одиннадцатый сегмент брюшка имеется у цикадовых и у самок скорпионниц.
В первом случае это анальная трубка, предотвращающая загрязнение тела испражняемым сиропом; она развита у цикадовых и редуцирована у полужесткокрылых [см. ниже, Hemelytrata (8) и рис. 9.7.1A]; о том, что это новообразование, свидетельствует тот факт, что у других Arthroidignatha, также обильно испражняющих сироп, подобного сегментоподобного образования нет.
Во втором случае речь идет об особой модификации брюшка самок, благодаря которой самка скорпионницы откладывает яйца в узкие укрытия, одновременно ощупывая эти укрытия вершиной брюшка, чтобы определить их влажность и пригодность для развития яиц. Авторы, считающие, что вершина брюшка самок скорпионниц несет одиннадцатый уромер и церки, вступают в противоречие, поскольку среди скорпионниц у самок Scorpiomuscae соции (принимаемые за церки) причленяются к анальному выступу (принимаемому за одиннадцатый уромер), а у Nannomecoptera соции причленяются к десятому уромеру по бокам от анального выступа [см. ниже, Scorpiomuscae (2), Nannomecoptera (5) и рис. 10.8.2].
В качестве вестигия одиннадцатого сегмента брюшка указывали непарный выступ между основаниями псевдокаудалий у двухвосток [см. ниже, Diplura (9)]. Однако этот выступ (эпипрокт) не несет на себе анальное отверстие, а находится дорсальнее его, так что он не является сегментом туловища. Псевдокаудалии не соединены с эпипроктом, а причленяются непосредственно к десятому сегменту брюшка.
(9) Трахейная система. Трахейная система [возникавшая у разных членистоногих независимо несколько раз – см. Gnathopoda (3c) и Atelocerata (6)], у Hexapoda имеет единое происхождение, о чем свидетельствует единое для Hexapoda расположение трахеостий.
(9 a) Строение трахейной системы. Трахеостии парные: на груди имеется до двух пар сегментарных трахеостий (мезоторакальные и метаторакальные) и до двух пар межсегментарных трахеостий (стеноторакальные и криптоторакальные), на брюшке имеются только сегментарные трахеостии [см. (4b)]. И сегментарные, и межсегментарные трахеостии обычно расположены на боках сегментов; лишь в тех случаях, когда форма сегмента изменена тем или иным образом, его трахеостии оказываются смещенными на дорсальную или на вентральную сторону. Обычно трахеостия находится между тергитом и стернитом – либо на плейральной мембране, либо на плейральном склерите (это может быть цельный плейрит или один из мелких склеритов). Иногда, благодаря разрастанию тергита или стернита на бока тела, трахеостии оказываются расположенными не между тергитом и стернитом, а на тергите (рис. 10.7.1) или на стерните (рис. 9.8.2A). Иногда расположение трахеостий не вполне соответствует внешней сегментации туловища: межсегментарная трахеостия может располагаться не только на межсегментной мембране, но и на склерите впереди лежащего сегмента или на склерите позади лежащего сегмента; cегментарная трахеостия может быть смещена на межсегментную мембрану или даже на склерит соседнего сегмента [см. ниже, Amyocerata (2)]. Иногда межсегментарные стеноторакальные дыхальца бывают так сильно смещены вперед, что их можно принять за сегментарные проторакальные дыхальца, которых на самом деле нет ни у каких насекомых [см. ниже, Mecopteriformia (5)]. В случаях увеличения заднегруди и редукции первого сегмента брюшка сегментарные дыхальца первого сегмента брюшка могут казаться расположенными на заднегруди; иногда они даже открываются непосредственно на плейрите заднегруди [см. ниже, Calyptroptera (3)].
Тогда как расположение трахеостий едино для Hexapoda, расположение отходящих от них трахей довольно разнообразно. Трахеи ветвятся. Пучки трахей, отходящих от разных трахеостий, могут быть независимыми, либо анастомозируют. Во многих случаях трахеи, идущие от всех трахеостий одной стороны тела, соединены общим продольным латеральным трахейным стволом; такой ствол имеется у части Diplura, Zygentoma и многих Pterygota. Кроме пары латеральных стволов, у некоторых насекомых имеются другие анастомозы, соединяющие трахеи разных трахеостий; иногда они образуют дополнительные парные продольные стволы – дорсальные, вентральные и/или висцеральные; иногда имеются поперечные вентральные и/или дорсальные анастомозы, соединяющие трахейные стволы левой и правой половины тела. Возможно, пара латеральных стволов является исходной для Hexapoda, а прочие стволы и анастомозы возникли независимо в разных группах. При линьке старая кутикула, выстилающая трахейный ствол, рвется на фрагменты по числу трахеостий, с которыми она связана, и каждый его фрагмент вместе со старой кутикулой трахей, отходящих от этого участка ствола, выходит через свою трахеостию.
Среди Hexapoda только у Collembola трахейная система не соответствует вышеописанной; судя по всему, у ногохвосток трахейная система исходно была утрачена, после чего заново возникла трахейная система иного строения, с дыхальцами на шее [см. ниже, Collembola (4)].
(9 b) Модификации трахейной системы. В большинстве случаев трахеостия представляет собой дыхальце – т.е. открываются наружу отверстием, через которое воздух попадает в трахеи; дыхальца могут иметь простое строение, либо снабжены фильтрующими и/или замыкательными аппаратами того или иного строения.
Иногда трахеостии определенных пар замкнуты и не функционируют. Нефункционирующая трахеостия снаружи может быть не выражена, но внутри от кутикулы отходит трахея [которая закладывается как впячивание кутикулы – см. Gnathopoda (3c)]; при линьке старая кутикулярная выстилка трахеи выходит через открытую трахеостию в новой неокрепшей кутикуле, после чего трахеостия замыкается и новая кутикула затвердевает так, что трахеостия остается постоянно замкнутой и не функционирует. Такие нефункционирующие трахеостии свойственны, в частности, некоторым личинкам, специализированным к обитанию в воде; в результате метаморфоза на месте нефункционирующих трахеостий появляются функционирующие дыхальца; таким образом, в процессе онтогенеза сохраняется один и тот же набор трахеостий, но может меняться их способность функционировать как дыхальца. Реже трахеостии определенных пар утрачены [см. ниже, Diplura (8), Protura (9), Amyocerata (2)].
У некоторых насекомых имеются вторичные дыхальца; в этом случае дыхальце находится не на самой трахеостии, а рядом с ней, и идущая к нему трахея отходит от трахеи, идущей от трахеостии; при этом трахеостия может быть замкнутой; при линьке старая кутикулярная выстилка всех трахей, включая дыхальцевую трахею, выходит не через вторичное дыхальце, а через трахеостию. Вторичные дыхальца имеются у многих личинок двукрылых [см. ниже, Diptera (4a)].
Некоторые насекомые вторично перешли к водному образу жизни и приспособились дышать растворенным в воде кислородом. В этом случае трахейная система сохраняется, так что дыхание осуществляется парадоксальным образом: растворенный в окружающей воде кислород через покровы насекомого диффундирует в трахеи, переходя в газообразное состояние, по трахеям разносится по телу, а в тканях опять переходит в водный раствор. У некоторых таких водных насекомых диффузия кислорода через покровы в трахеи происходит преимущественно в выступающих тонкостенных придатках – трахейных жабрах (branchiae tracheales); в разных таксонах в качестве трахейных жабр служат те или иные придатки – максиллярные и/или лабиальные щупики, тергалии [см. ниже, Ephemeroptera (12)], эпипрокт и/или парапрокты [см. ниже, Zygoptera (1)], либо новообразования, независимо возникшие у разных насекомых на тех или иных частях тела. У многих из этих водных насекомых трахейная система замкнутая, то есть трахеостии постоянно закрыты и не служат дыхальцами; в некоторых случаях одновременно имеются и трахейные жабры, и функционирующие дыхальца. У некоторых водных насекомых брюшко несет латеральные отростки, которые можно назвать придыхальцевыми трахейными жабрами: трахея, входящая в такой отросток, начинается непосредственно от трахеи, идущей от дыхальца того же сегмента брюшка (подобно трахеи, идущей к вторичному дыхальцу – см. выше); сходные по строению придыхальцевые трахейные жабры независимо друг от друга возникли у некоторых личинок крылатых насекомых [см. ниже, Metapterygota (6)]. Многие водные насекомые являются амфибиотическими: их преимагинальные стадии обитают в воде, а имаго на суше. Подводные и амфибиотические представители имеются только среди Pterygota (см. Указатель признаков [6.1]), тогда как среди первичнобескрылых насекомых (Entognatha и Triplura) подводных форм нет. Подавляющее большинство водных насекомых обитают только в пресных водах, лишь единичные виды приспособились к морской среде (Cheng 1976) [см. ниже, Diptera (4) и Trichoptera (1)].
Высказывалось предположение, что предки Pterygota перешли к жизни в воде, после чего имаго вторично вышли на сушу, и, таким образом, водный образ жизни личинок поденок, стрекоз, веснянок, вислокрылок и других амфибиотических насекомых является не апоморфией, а плезиоморфией среди Pterygota. В качестве аргумента в пользу этой точки зрения приводили тот факт, что имаго некоторых видов имеют вестигиальные трахейные жабры в виде небольших мягких отростков. В действительности у всех имаго, которые имеют жаберные отростки, эти отростки представляют собой не остатки имагинальных жабр древних предков, а остатки личиночных жабр той же особи, которые не до конца резорбировались при превращении личинки в имаго [см. ниже, Ephemeroptera (1d), Odonata (1a), Plecoptera (1D), Meganeuroptera (1)].
(9 c) Сравнение. Помимо Hexapoda, трахеостии на плейритах туловища имеются только у губоногих многоножек (рис. 5.5.3), но там они иначе распределены по сегментам [ср.: Pleurostigmophora (7) и рис. 5.5.4]; у этих многоножек имеются также продольные трахейные стволы, сходные с латеральными стволами насекомых. Судя по тому, что в голофилетическом таксоне Chilopoda трахеостии по-разному расположены у Pleurostigmophora и Notostigmophora, сходное расположение трахеостий у Pleurostigmophora и Hexapoda возникло независимо.
(10) Прилежащие тела. Имеются прилежащие тела (corpora allata) – пара эндокринных желез эктодермального происхождения, лежащих по бокам аорты и относящихся к мандибулярному или максиллярному сегменту; единое происхождение прилежащих тел подтверждается их одинаковой иннервацией. Прилежащие тела секретируют ювенильный гормон (hormonum juvenile) играющий важную роль в регуляции развития. Прилежащие тела имеются как у Amyocerata, так и у Entognatha, и принципиально отличаются от секреторных органов, имеющихся у Myriapoda (Sahli 1985).
Признаки Hexapoda неясного филогенетического значения
(11) Сегментарный состав головы. Головная капсула, или краниум (cranium) состоит из полностью слившихся преоральной области и I–IV посторальных сегментов [см. Metameria (1)]; таким образом, голова несет одну пару антенн [преоральных – см. Atelocerata (1)] и три пары специализированных ротовых конечностей – мандибулы [см. Mandibulata (1)], максиллы [см. (2)] и нижнюю губу [см. (3)]; следующие сегменты (начиная с V посторального) с головой не сливаются и образуют обособленную от головы грудь [см. (4)]. Такой же сегментарный состав слитной головной капсулы имеется у Symphyla, Chilopoda и некоторых Eucrustacea, включая такие примитивные группы, как Cephalocarida. Возможно, этот сегментарный состав головы исходно свойственен Mandibulata. В то же время у некоторых других Mandibulata сегментарный состав головы иной: у Collifera в состав головной капсулы не входит IV посторальный сегмент; у некоторых ракообразных протоцефалон и гнатоцефалон не слиты; у некоторых ракообразных, наоборот, в состав слитной головной капсулы входит большее число сегментов (ср.: Eucrustacea: «Классификации» II). В отличие от Hexapoda, у некоторых других членистоногих специализация конечностей не всегда коррелирует с их принадлежностью к голове: например, у Cephalocarida, а также у Trilobita некоторые конечности, находящиеся на слитной голове, не отличаются от грудных (рис. 4.1). Можно предположить, что слияние головных сегментов происходило независимо в разных группах Mandibulata, и данное строение головы может быть аутапоморфией Hexapoda.
(12) Строение гонад. Гонады, особенно у самок, имеют некоторые особые черты строения. Гонады самцов и самок [обычно парные, образованные мезодермальным эпителием – см. Gnathopoda (1)] ориентированы так, что их выводные протоки направлены назад и открываются в задней части тела (в отличие от Progoneata).
Положение половых отверстий несколько различается в разных группах Hexapoda: у Entognatha оно исходно находится между VIII и IX сегментами брюшка [см. ниже, Entognatha (4)]; у самцов Amyocerata – между IX и X сегментами брюшка [см. ниже, Amyocerata (6)]. У самок различных представителей Amyocerata половое отверстие открывается в разных местах – от границы VII и VIII сегментов до IX сегмента (см. Указатель признаков [4.11.8]); вероятно, его смещение объясняется редукцией первичного яйцеклада [см. (13)] и образованием на его месте вторичных стернитов. По-видимому, первичным для Amyocerata является положение полового отверстия самки на границе VII и VIII сегментов брюшка. Об этом свидетельствует то, что при различных положениях полового отверстия парные протоки, идущие к нему от гонад, обходят вентральную нервную цепочку между нервными стволами VII и VIII уромеров (Brinck 1956).
По всей видимости, исходным для Hexapoda является непарное половое отверстие и у самца, и у самки: пара мезодермальных половых протоков кзади сливается в непарный эктодермальный проток, идущий вентральнее кишечника и открывающийся непарным половым отверстием. У самцов некоторых насекомых (Protura, большинство Ephemeroptera, некоторые Plecoptera, некоторые Dermatoptera, некоторые Auchenorrhyncha и др.) половые отверстия парные; при этом в одних случаях непарный эктодермальный проток отсутствует, а в других случаях непарный половой проток имеется, но дистально он ветвится и открывается парой отверстий; вероятно, и то, и другое является вторичными преобразованиями [см. ниже, Ephemeroptera (15a), Plecoptera (10), Dermatoptera (9), Hemelytrata (7)]; у некоторых двукрылых число половых отверстий самца увеличено до 3, а у некоторых уховерток – до 4 [см. ниже, Diptera (5) и Dermatoptera (9)].
Гонады самок (яичники, или оварии) имеют следующие особенности. У большинства (но не у всех) Hexapoda каждый из пары яичников (ovarium) состоит из нескольких овариол, выводные протоки которых соединены в общий яйцевод. Каждая овариола (ovariolum) представляет собой трубку, внутри которой созревающие яйцеклетки расположены в один ряд; каждой яйцеклетке соответствуют вздутие овариолы – фолликул (folliculus). Апикальная часть овариолы (т.е. ее передняя часть по отношению к оси тела), называемая гермарий (germarium), наиболее узкая, в ней находятся беспорядочно расположенные оогонии и ооциты на ранних стадиях созревания [см. Metazoa (2)]; по мере своего созревания яйцеклетки движутся по овариоле друг за другом, так что наиболее зрелые яйца, находящиеся в наиболее крупных фолликулах овариолы, расположены в проксимальной (т.е. задней) части овариолы, называемой вителлярий (vitellarium).
У некоторых Hexapoda (в частности, у Diplura-Dicellurata и Microcoryphia) в каждом яичнике имеется 7 овариол, гребенчато впадающих в общий яйцевод – т.е. число пар овариол соответствует числу сегментов брюшка, в которых помещаются яичники. В связи с этим некоторые авторы интерпретируют овариолы как исходно сегментарные органы, и считают, что такое строение яичника, когда он состоит из овариол, является плезиоморфией. Однако у разных насекомых число пар овариол различно, оно бывает значительно меньшим или значительно большим числа сегментов – от одной пары до нескольких сотен; часто овариолы впадают в общий проток не гребенчато, а пучком, так что их сегментарная природа не очевидна. У других членистоногих яичник не разделен на овариолы, поэтому можно предположить, что разделение яичника на овариолы является аутапоморфией Hexapoda. У некоторых Hexapoda каждый из пары яичников состоит лишь из одной овариолы либо не разделен на овариолы. В связи с этим высказано предположение, что наличие овариол является апоморфией не Hexapoda в целом, а меньшего таксона под названием Ovariolata Büning 1996, объединяющего Dicellura и Amyocerata, но не включающего Rhabdura и Ellipura (у которых яичник состоит из пары цельных овариев, не разделенных на овариолы). Однако выделение таксона Ovariolata противоречит представлению о голофилии Entognatha и голофилии Diplura. Яичник, вторично утративший разделение на овариолы, известен для некоторых Diptera и некоторых Hymenoptera. Вероятно, отсутствие овароил у Rhabdura и Ellipura также является вторичным.
У насекомых принято различать паноистические яичники (когда нет специальных питающих клеток) и мероистические яичники, когда помимо ооцитов имеются особые питающие клетки – трофоциты (trophocyti). Во втором случае каждый ооцит соединен с одним или многими трофоцитами цитоплазматическими мостиками. Трофоциты и ооциты (называемые общим термином цистоциты – cystocyti) получаются в результате следующих друг за другом митотических делений одной предшествующей им клетки (оогония, или цистобласта – cystoblastus) и образуют кластер из клеток, исходно соединенных цитоплазматическими мостиками. Мероистические яичники бывают политрофическими и телотрофическими. В политрофическом яичнике все трофоциты, связанные с яйцеклеткой, находятся в том же фолликуле, что и эта яйцеклетка. При этом обычно (у Panpsocoptera и многих Metabola) в кластере, получившимся в ходе многократного деления одного цистобласта, лишь одна клетка развивается в ооцит, а остальные становятся трофоцитами; так что число трофоцитов в каждом фолликуле обычно оказывается равным 2n–1 (1, 7, 15 и т.д., до 255), но иногда развивается иное число трофоцитов (меньше или больше исходного 2n–1). В телотрофическом (или акротрофическом) яичнике трофоциты всех яйцеклеток, имеющихся в одной овариоле, сконцентрированы в апикальной части овариолы, называемой трофарий (tropharium); от каждой яйцеклетки к трофарию тянется длинный цитоплазматический шнур, соединяющий яйцеклетку с трофоцитами. При этом в кластере, получившимся из одного цистобласта, может быть несколько яйцеклеток; трофоциты в трофарии бывают соединены в один синцитий и в этом случае являются общими для всех яйцеклеток данной овариолы. Принято считать, что исходным для Hexapoda является паноистический тип яичника, а политрофический и телотрофический типы характерны для отдельных таксонов в составе Hexapoda (см. Указатель признаков [4.11.4]); оба эти типа возникали в ходе эволюции по нескольку раз; судя по всему, несколько раз происходил и обратный переход к паноистическому типу (Bonhag 1958, Büning 1993, 1994, 1996). Иногда вторично возникший паноистический тип называют неопаноистическим, хотя он ничем не отличается от первичного паноистического.
Среди Diplura паноистические яичники описаны у Dicellurata, политрофические – у Rhabdura. У Collembola яичники политрофические, у Protura – паноистические . У Triplura яичники паноистические. У яичников Ephemeroptera обнаружено особое телотрофическое строение (Gottanka & Büning 1993). У Odonata, Embioptera, Notoptera, Plecoptera, Pandictyoptera, Saltatoria и Spectra яичники паноистические. У уховерток яичники политрофические [см. ниже, Dermatoptera (12)]. Среди Parametabola и Metabola наиболее обычны политрофические и телотрофические яичники, но встречаются и паноистические [см. ниже, Eumetabola (6), Arthroidignatha (4), Calyptroptera (6)].
И у самца, и у самки позади гонад обычно имеется пара придаточных желез (glandulae accessoriae), протоки которых открываются либо позади гонопора, либо в эктодермальный гонодукт. У самцов секрет придаточных желез либо выводится в виде жидкости вместе со сперматозоидами, либо затвердевает, образуя вокруг сперматозоидов капсулу – сперматофор (spermatophorus). У самок секрет придаточных желез может служить для склеивания откладываемых яиц или приклеивания их к субстрату; в этом случае придаточные железы называют коллетериальными (colleteria).
(13) Яйцеклад. Вероятно, самки Hexapoda исходно имеют яйцеклад, образованный модифицированными кинетапофизами VIII и IX пар [см. (7)]. Судя по тому, что такой яйцеклад есть у всех Triplura и части Pterygota, он, бесспорно, существовал у общего предка Amyocerata [см. ниже, Amyocerata (7)]. Ни у одного известного представителя Entognatha яйцеклада нет. Однако у Entognatha-Diplura кинетапофизы и стерностили бывают развиты только на I–VII уромерах и всегда отсутствуют на VIII–IX уромерах [см. ниже, Diplura (7)]. Это дает основание предполагать, что именно на этих сегментах брюшка кинетапофизы были исходно преобразованы в створки яйцеклада, а затем редуцировались (подобно тому, как яйцеклад редуцировался у многих Pterygota).
(14) Форма линочного шва. Линочный шов, по которому при линьке трескается старая кутикула [см. Gnathopoda (3b)], обычно имеет следующую форму (рис. 7.2.3С): на головной капсуле имеется пара лобных (или фронтальных) швов (suturae frontales), которые кзади сходятся в один непарный продольный дорсальный шов, идущий далее назад медиально по голове и по тергитам груди. При наличии трех глазков [см. ниже, Amyocerata (4)] парные лобные швы проходят позади медиального глазка, а их передние концы могут оканчиваться вблизи парных глазков. Область головы впереди и медиальнее лобных швов принято называть лоб (frons). Такая форма линочного шва свойственна подавляющему большинству преимагинальных возрастов насекомых. Лишь в немногих случаях форма линочного шва изменена (см. Указатель признаков [5.8]); у некоторых стадий кутикула настолько тонкая, что при линьке рвется в произвольных местах.
У других членистоногих формы линочного шва иные. У Chilopoda имеется лишь поперечный шов на голове и нет продольного шва. У Eucrustacea-Malacostracia кутикула распадается на переднюю и заднюю части по кольцевому шву, опоясывающему туловище; в случае наличия карапакса этот шов отделяет несегментированный карапакс от сегментированной части туловища; в случае отсутствия карапакса он разделяет две сегментированные части туловища (мною наблюдались представители Decapoda и Isopoda-Terrestria). У Chelicerata-Maripalpi имеется кольцевой шов, отделяющий тергит головогруди от остальной кутикулы. У Trilobita линочные швы находятся в пределах нерасчлененного головного отдела (включающего три посторальных сегмента – см. Trilobitomorpha: «Классификация») – это пара лицевых, или фациальных, швов, проходящих по дорсальной стороне головы и отделяющих медиальную часть головы с глазами (cranidium) от ее латеральных частей (librigenae); непарный шов отсутствует.
Геологический возраст. Девон – ныне (о древнейшем известном представителе – см. ниже, Collembola).
Систематическое положение Hexapoda. Помимо принятого здесь отнесения Hexapoda непосредственно к Atelocerata, предлагались следующие классификации.
I. Объединение Hexapoda, Symphyla и Chilopoda в таксон Etymachila (см. Atelocerata: «Классификации» I).
II. Объединение Hexapoda и Symphyla в таксон Dimalata (см. Atelocerata: «Классификации» II).
III. Объединение Hexapoda и Progoneata в таксон Labiata (см. Atelocerata: «Классификации» III).
IV. Объединение Hexapoda и Chilopoda в таксон Opisthogoneata (см. Atelocerata: «Классификации» V).
V. Объединение Hexapoda и Eucrustacea в таксон Pancrustacea (см. Mandibulata: «Классификации» II).
Статус Hexapoda. В некоторых классификациях таксон Hexapoda отсутствует (см. Atelocerata: «Классификации» IV). В литературе (Sharov 1966, Расницын 1980 и др.) фигурирует утверждение, будто бы существует обоснованная и широко признанная теория о полифилетическом происхождении Hexapoda, в связи с чем некоторые авторы считают нецелесообразным выделение этого таксона. В результате, во многих публикациях таксоны Amyocerata (под названием «Ectognatha» или «Insecta») и Entognatha рассматриваются не в составе Hexapoda, а как не связанные друг с другом таксоны в составе Atelocerata. В действительности никто никогда не выдвигал и не обосновывал теорию о полифилии Hexapoda, а недоразумение вызвано тем, что нетипифицированное название Insecta используется в качестве рангового (см. раздел I.6.6.2). Обоснованием теории о полифилии Hexapoda могли бы быть какие-нибудь признаки, считающиеся апоморфиями и являющиеся общими для какой-либо отдельной группы внутри Hexapoda и какой-либо группы животных, не входящих в Hexapoda (например, каких-либо Myriapoda). Однако такие апоморфии не описаны.
Говоря о полифилии Hexapoda, иногда ссылаются на теорию Берлезе (предполагавшую особое положение Protura) и теорию Тиллиярда (предполагавшую особое положение Collembola); в действительности в обеих этих теориях говорится не о полифилии Hexapoda, а лишь о полифилии Entognatha (см. ниже, Collembola: «Систематическое положение» II; Protura: «Систематическое положение» I).
Некоторые авторы активно отстаивают идею о полифилии Hexapoda. В качестве довода в пользу полифилии Hexapoda они утверждают, что им неизвестно никаких других апоморфий Hexapoda, кроме шестиногости, а шестиногость не является абсолютно надежной апоморфией и может возникать у разных членистоногих [см. Hexapoda (4c)]. Даже если бы шестиногость была единственным аргументом в пользу голофилии Hexapoda, не было бы повода говорить о полифилии Hexapoda, так как в пользу полифилии не названо ни одного аргумента. Однако шестиногость является далеко не единственной апоморфией Hexapoda. Прочие апоморфии Hexapoda подробно описаны во всех учебниках энтомологии; однако из большинства публикаций, где описывается строение так называемых «Insecta», невозможно понять, какой именно таксон в данном случае подразумевается – Hexapoda, Amyocerata или какой-нибудь иной (см. раздел I.6.6.2).
Классификации Hexapoda.
I. Деление Hexapoda на Apterygota и Pterygota. Долгое время было принято деление Hexapoda на первичнобескрылых (Apterygota) и крылатых (Pterygota). Таксон Apterygota Lang 1888 – первичнобескрылые {synn.circ.: Thysanoura Latreille 1796 = Seticaudes Cuvier 1805 = Nematoura Dumeril 1805 = Thysanura Leach 1815a = Thysanuria Rafinesque 1815 = Monomorphes Lucas 1840 = Apterygogenea Brauer 1885 = Apterota Haeckel 1896 = Aptilota Lameere 1935 = Epimetabola Weber 1933 (название для типа метаморфоза)} объединяет Entognatha и Triplura. Он является парафилетическим и предковым для голофилетического таксона Pterygota. Высказывалась также альтернативная точка зрения, что Apterygota произошли от Pterygota в результате редукции крыльев (Handlirsch 1908); против такой точки зрения свидетельствуют апоморфии Amyocerata (см. ниже) и апоморфии Pterygota (см. ниже).
II. Деление Hexapoda на Entognatha и Amyocerata. Эта классификация основана на представлении о том, что Triplura имеют общего предка с Pterygota. На основании этого Triplura и Pterygota объединяются в голофилетический таксон Amyocerata, тогда как Diplura, Collembola и Protura объединяются в другой голофилетический таксон – Entognatha (об апоморфиях Entognatha и Amyocerata см. ниже). При этом таксон Apterygota не признается, поскольку является парафилетическим; таксон Pterygota в этой классификации оказывается подчиненным в составе Amyocerata. В данной книге мы придерживаемся именно этой классификации:
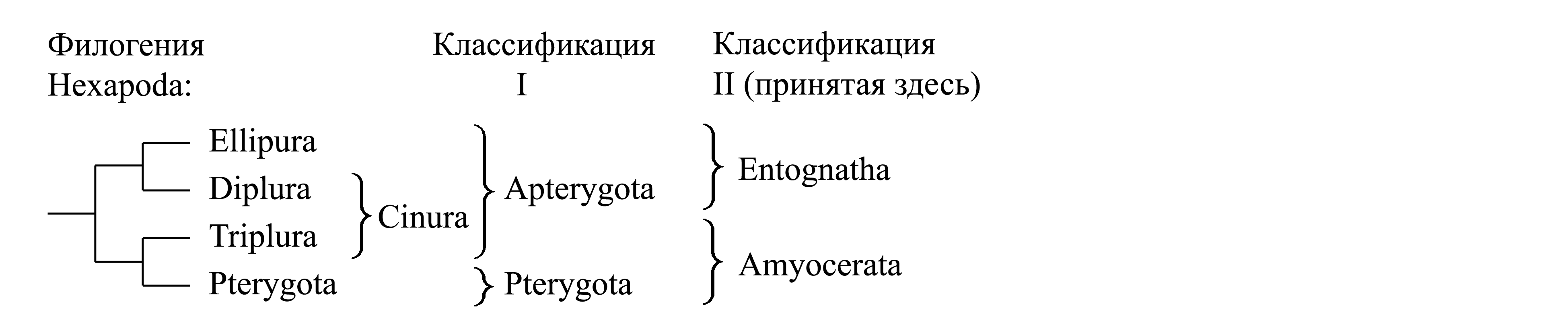
III. Деление Hexapoda на Ellipura и Pleomerentoma. Эта классификация основана на иной филогенетической гипотезе:

Основанием для этой гипотезы служит сходство в строении брюшка Diplura и Triplura [см. Hexapoda (7)], которых вместе с Pterygota объединяют в таксон Pleomerentoma Krausse & Wolff 1919 {synn.circ.: Euentomata Imms 1936 = Epimorpha Chen 1962 (non Epimorpha Haasae 1880) = Cercophora Willmann 2003 (non Cercophora Börner 1904); по циркумскрипции также соответствует: «Insecta» sensu Chen 1962 (non Linnaeus 1758)}. Предполагаемой апоморфией Pleomerentoma является то, что проксимальные членики редуцированных абдоминальных конечностей вошли в состав вентральной стенки сегмента брюшка, так что втяжные пузырьки оказались сидящими прямо на брюшке (рис. 6.6.3); у Ellipura втяжные пузырьки сидят на вершинах предполагаемых вестигиев абдоминальных конечностей, что рассматривалось как плезиоморфия (Kukalova-Peck 1987, 1991) (о сомнениях на этот счет см. ниже, Ellipura: «Систематическое положение»). Эта теория противоречит представлению о голофилии Entognatha, которая доказывается уникальным строением головы [см. ниже, Entognatha (1)]. Судя по тому, что Diplura и Ellipura образуют голофилетический таксон Entognatha, общие признаки Diplura и Triplura являются не синапоморфиями, а симплезиоморфиями.
IV. Деление Hexapoda на Protura и Holomerentoma (см. ниже, Protura: «Систематическое положение» I).
V. Деление Hexapoda на Protura, Collembola и Pleomerentoma (см. III и ниже, Collembola: «Систематическое положение» II).
Здесь мы принимаем деление Hexapoda на два подчиненных таксона – Entognatha и Amyocerata (с. 0000).
Hexapoda, или Pediculus/f3=Scarabaeus/g3
![]() 1. Entognatha, или Podura/fg1
1. Entognatha, или Podura/fg1
![]() 1.1. Diplura, или Iapyx/f1=Campodea/g1
1.1. Diplura, или Iapyx/f1=Campodea/g1
![]() 1.2. Ellipura, или Podura/fg2
1.2. Ellipura, или Podura/fg2
![]() 1.2.1. Collembola, или Podura/fg3
1.2.1. Collembola, или Podura/fg3
![]() 1.2.2. Protura, или
Acerentomon/fg1
1.2.2. Protura, или
Acerentomon/fg1
![]() 2. Amyocerata, или Pediculus/f4=Scarabaeus/g4
2. Amyocerata, или Pediculus/f4=Scarabaeus/g4
![]() 2.1. Triplura, или Lepisma/fg1
2.1. Triplura, или Lepisma/fg1
![]() 2.1.1. Zygentoma, или Lepisma/fg2
2.1.1. Zygentoma, или Lepisma/fg2
![]() 2.1.2. Microcoryphia, или
Machilis/fg1
2.1.2. Microcoryphia, или
Machilis/fg1
![]() 2.2. Pterygota, или Pediculus/f5=Scarabaeus/g5
2.2. Pterygota, или Pediculus/f5=Scarabaeus/g5