КЛАДОЭНДЕЗИС НАСЕКОМЫХ

![]()
|
КЛАДОЭНДЕЗИС НАСЕКОМЫХ |
|||||||
|
|
|
fg | |||||
Глава IV. Общая система членистоногих
IV-1. Gnathopoda Lankester 1877 – членистоногие в широком смысле
(рис. 4.1–10.16)
|
Названия, однозначно соответствующие по циркумскрипции: — Gnathopoda Lankester 1877 (non Gnathopodaria Pictet 1854); — Lobopoda Boudreaux 1979a; — Panarthropoda Nielsen 1995; — Podophora Waggoner 1996; — Antennopoda De Haro 1998; — Haemopoda Cavalier-Smith 1998. Названия, неоднозначно соответствующие по циркумскрипции (соответствующие также Euarthropoda) – см. ниже, Euarthropoda. По циркумскрипции также соответствует: — «Arthropoda» auct. (non Siebold 1848). Иерархическое название: Araneus/fg6 [fg:(1757)1758] (sine Lumbricus; incl. Peripatus, Macrobiotus). |
|
Ранг. Обычно тип или подтип.
Название. Авторство названия Lobopoda, или Lobopodia, приписывают Снодгассу (Snodgrass 1938a), который использовал лишь английское словосочетание «lobopod branch» в филогенетической схеме (Snodgrass 1938a: Fig. 54); здесь слово «lobopod» использовано не в качестве названия таксона, а в качестве эпитета, означающего, что в данной ветви конечности приобрели вид лобоподий; из текста и схемы неясно, относится ли этот термин ко всей ветви, включающей Onychophora и Euarthropoda, или только к предковой группе в основании этой ветви. В последствие некоторые авторы (Boudreaux 1979a, Клюге 1999, 2000) приписывали название «Lobopoda Snodgrass 1938» таксону, объединяющему Onychophora с Euarthropoda – т.е. членистоногим в широком смысле слова, тогда как другие авторы (Hou & Bergstrom 1995) приписывали название «Lobopodia Snodgrass 1938» небольшой группы животных, не включающей Euarthropoda. Во избежание путаницы следует использовать наиболее старое название Gnathopoda, которое было предложено для членистоногих в широком понимании, включая Onychophora. По своей этимологии слово «Gnathopoda» («челюстноногие») не соответствует характеристике данного таксона: хотя у Onychophora и многих Euarthropoda передние пары конечностей действительно специализировались как челюсти, у тихоходок челюсти не гомологичны ногам [см. ниже, Tardigrada (1)]; вероятно, те или иные конечности специализировались как челюсти независимо в разных группах Gnathopoda, и общий предок Gnathopoda не имел челюстей, гомологичных конечностям. Однако этимология не должна влиять на выбор названия (см. I.6.2).
Иногда этому таксону ошибочно приписывают название «Arthropoda»; в действительности название Arthropoda Siebold 1848 исходно было дано таксону, заведомо не включавшему онихофор, которые в той же книге рассматривались как род Peripatus в составе кольчатых червей (Siebold 1848) (см. «Классификации» II).
Название Gnathopoda сходно с более старым названием Gnathopodaria, но не идентично ему; название Gnathopodaria Pictet 1854 (только во французском написании, как «Gnathopodaires»), было дано для таксона, объединяющего всех членистоногих, кроме Hexapoda.
Аутапоморфии Gnathopoda
(1) Редукция целома. Целомические мешки [исходно парные, сегментарные – см. Metameria (1)] редуцированы по крайней мере у взрослых животных; так что полость тела представляет собой миксоцель (mixocoelom) – результат объединения целомической полости [см. Peritoneata (1)] и первичной полости тела (бластоцеля). Редукция целома не связана с какими-либо адаптациями [как и появление целома у общего предка целомических животных – см. Peritoneata (1)]. Вестигии целома (исходно выполнявего и половую, и выделительную функции) преобразованы в разобщенные органы – выделительные железы и гонады.
Выделительные железы парные, каждая железа представляет собой мезодермальный целомический мешок с эктодермальным выводным протоком – целомодуктом. У Onychophora по паре таких желез развито во многих сегментах тела (рис. 4.2D); у части Chelicerata и Eucrustacea бывает развита одна или несколько пар выделительных желез (см. Указатель признаков [4.7.1]). У Tardigrada, Pantopoda и некоторых других эти железы утрачены. У Atelocerata в связи с развитием выделительных мальпигиевых сосудов эти железы утратили выделительную функцию и, вероятно, преобразовались в некоторые из слюнных, паутинных и других желез, открывающихся в ротовом аппарате (см. Указатель признаков [4.3.6–9]).
В то время как у многих животных, обладающих целомом, половые продукты формируются на внутренней стенке целома и попадают в целомическую полость [см. Peritoneata (1)], у Gnathopoda имеются эпителизованные гонады (gonadae) – семенники (testes) у самца и яичники (ovaria) у самки, из которых половые продукты выводятся по специальным протокам – гонодуктам (gonoducti) [соответственно, семяпроводам (vasa deferentia) у самца и яйцеводам (oviducti) у самки], открывающимся наружу отверстиями – гонопорами (gonopori). В некоторых случаях гонодукты явно гомологичны целомодуктам: в частности у Onychophora на сегменте, несущем гонопор, нет целомодуктов, тогда как на других сегментах они развиты. Однако, судя по тому, что у разных членистоногих гонодукты открываются на разных сегментах (см. Указатель признаков [4.11.7]), во многих случаях гонодукты возникали вторично как новообразования [в частности, см. ниже, Progoneata (1)].
Поскольку целомические мешки исходно парные [см. Peritoneata (1)], то гонады, гонодукты и гонопоры также исходно должны быть парными. Но у некоторых членистоногих они становятся непарными: левая и правая гонады бывают частично или полностью слиты между собой дорсальнее или вентральнее кишечника, но всегда дорсальнее нервной цепочки [см. Metameria (2)]; часто парные половые протоки впадают в непарный проток, который открывается непарным гонопором. Если гонопор непарный, он всегда находится на вентральной стороне тела впереди анального отверстия; соответственно в этом случае парные гонодукты обходят с боков нервную цепочку и сливаются в непарный проток вентральнее нервной цепочки. Некоторые авторы считают, что эволюция во всех случаях шла только от парных половых органов к непарным; однако, судя по всему, во многих случаях происходил также и переход от непарных органов к парным (см. Указатель признаков [4.11.6]).
В эмбриогенезе у многих членистоногих во всех или в некоторых сегментах закладываются целомические мешки, но затем они распадаются; гонады и выделительные железы развиваются либо из фрагментов эмбриональных целомических мешков, либо возникают заново после распада целомических мешков; у некоторых членистоногих целомические мешки в эмбриогенезе вообще не закладываются. Эмбриональные метамерные парные целомические мешки могут закладываться не только в посторальных сегментах (по паре в каждом сегменте), но и в преоральной области [см. Metameria (2)].
(2) Незамкнутая кровеносная система. Кровеносная система [см. Metameria (4)] незамкнутая, т.е. сосуды открываются в миксоцель [см. (1)]; из миксоцеля кровь засасывается обратно в продольный дорсальный сосуд через имеющиеся в нем остии (ostia) – отверстия, снабженные клапанами. В результате этого в сосудах и в миксоцеле находится единая жидкость – гемолимфа (haemolympha). Поскольку продольный дорсальный сосуд на всем своем протяжении или на определенном участке снабжен мышцами и способен к ритмическим сокращениям, его принято называть сердцем. У большинства Gnathopoda кровеносные сосуды подвергаются значительной редукции: иногда сохраняются оба продольных сосуда (дорсальный и вентральный) и отходящие от них сегментарные парные сосуды; часто вентральный сосуд редуцируется и остается только дорсальный сосуд – сердце [по которому кровь гонится преимущественно вперед – см. Metameria (4)]; у некоторых мелких Gnathopoda кровеносная система совсем исчезла (в отличие от Annelida, у которых замкнутая кровеносная система сохраняется даже при малых размерах животного).
Среди Gnathopoda наиболее полное строение кровеносной системы сохраняется у Chilopoda: у них имеется не только дорсальный сосуд (сердце), но и вентральный кровеносный сосуд, который в передней части соединен с дорсальным сосудом посредством артериального кольца, огибающего кишку; имеются также разветвленные сосуды, отходящие как от дорсального, так и от вентрального сосуда и открывающиеся в миксоцель. Помимо Chilopoda, сложное строение кровеносной системы свойственно некоторым Chelicerata и некоторым Malacostracia. Сильно упрощенное строение с одним лишь дорсальным сосудом свойственно таким далеким друг от друга группам, как Onychophora, некоторые Chelicerata, некоторые Eucrustacea, Progoneata и Hexapoda. Кровеносная система полностью утрачена у Tardigrada и у наиболее мелких представителей Arachnida и Eucrustacea. Из этого следует, что у Gnathopoda происходило неоднократное упрощение кровеносной системы и, возможно, неоднократное ее усложнение, но при этом она всегда остается незамкнутой.
Помимо Gnathopoda, среди Metameria кровеносная система редуцирована у челюстных пиявок (Gnathobdellea), но там это связано с тем, что целом превратился в систему каналов и функционально заменил кровеносную систему. Незамкнутая кровеносная система встречается также у некоторых животных, не относящихся к Metameria – это иглокожие (Echinodermata), некоторые щупальцевые (Lophophorata), некоторые моллюски (Malacozoa).
Было высказано предположение, что редукция кровеносной системы у Gnathopoda связана с редукцией целома [см. (1)], поскольку у Annelida кровеносные сосуды, будучи зажатыми между целомическими мешками, в значительной степени образованы за счет материала целомического эпителия (Беклемишев 1964); если это так, то незамкнутость кровеносной системы и редукцию целома следует рассматривать как один признак, а не как две разные аутапоморфии. Однако среди разных групп животных нет корреляции между наличием целома и замкнутостью кровеносной системы.
(3) Кутикула и линьки. Имеется хитинсодержащая внеклеточная кутикула [см. (3a)], в связи с этим происходят регулярные линьки [см. (3b)]. Кутикула развита на эпителии эктодермального происхождения [см. Monostomata (1)], так что она покрывает все животное снаружи и выстилает изнутри переднюю и заднюю кишки [см. Monostomata (2) и Haemataria (1)] [см. (3d)]. В связи с развитием кутикулы эктодермальный эпителий полностью утратил подвижные реснички [см. Monostomata (1)], в нем сохраняются лишь сенсорные реснички [см. Monostomata (3b)]; соответственно, утрачена и стадия трохофоры [см. Metameria (6)]. Вероятно, наличие кутикулы привело тому, что в разных группах членистоногих независимо несколько раз возникала трахейная система, выстланная кутикулой [см. (3c)].
Хитинсодержащая кутикула, сбрасываемая при линьках, свойственна всем Gnathopoda – Onychopohora (Storch & Ruhberg 1993), Tardigrada (Dewel, Nelson & Dewel 1993) и настоящим членистоногим [см. ниже, Euarthropoda (1)]. Способность вырабатывать хитин исходно присуща животным [см. Eucaryota (5)], так что аутапоморфией Gnathopoda является не наличие хитина, а наличие хитинсодержащей внеклеточной кутикулы. По строению кутикулы Gnathopoda отличаются от Annelida и большинства других животных [см. (3e)].
Помимо кутикулы, эктодерма членистоногих может выделять другие хитинсодержащие внеклеточные образования, такие как перитрофическая мембрана [см. (3d)] и выстилка воздухоносных полостей в легких у паукообразных [см. ниже, Chelicerata: «Классификации» III: Arachnida (5b)].
(3 a) Строение кутикулы. Покровный эпителий состоит из одного слоя клеток и покрыт снаружи сплошным слоем кутикулы (cuticula), выделяемой эпителиальными клетками [в связи с тем, что эпителий находится под кутикулой, его принято называть гиподерма (hypoderma)]. Кутикулярные структуры могут быть выявлены при обработке животного щелочью: в то время, как все ткани в щелочи растворяются, кутикула сохраняется. Кутикула состоит из хитина [см. Eucaryota (5)] и различных белков; белок коллаген не входит в состав кутикулы, хотя имеется в организме и участвует в образовании внутренних соединительных тканей (в частности, у Onychophora, имеющих тонкую кутикулу, под гиподермой имеется толстый слой ткани, содержащей коллаген). Характерным для кутикулы Gnathopoda считается наличие α-хитина (Storch & Ruhberg 1993), но при этом организм может вырабатывать также β-xитин и γ-хитин (Rudal & Kenchington 1973). Кутикула, состоящая только из хитина, представляет собой прочную, бесцветную, прозрачную эластичную пленку; благодаря наличию тех или иных белков, связанных с хитином, определенные участки кутикулы окрашены и склеротизованы [в частности, склериты настоящих членистоногих – см. ниже, Euarthropoda (1)], тогда как другие участки сохраняют гибкость. Кутикула имеет сложное слоистое строение; разные слои могут различаться концентрацией и ориентацией волокон хитина, составом белков и других веществ; наружный слой, называемый эпикутикула (epicuticula), может не содержать хитин и состоять только из белков и липидов. Подробнее см. Тыщенко 1976.
У части наземных членистоногих кутикула, помимо хитиново-белковых и белковых слоев, снаружи имеет еще один слой – восковой; химический состав воска сильно различается у разных видов. Воск выделяется железами, расположенными в гиподерме, и, проходя сквозь кутикулу, изливается на ее поверхность; благодаря этому обеспечивается защита животного от высыхания, а поверхность кутикулы становится гидрофобной (подробнее см. Lower 1959). Такой восковой покров имеется в далеких друг от друга таксонах членистоногих – у наземных Hexapoda и Arachnida; у водных и некоторых наземных членистоногих восковой покров отсутствует.
(3 b) Линьки. Поскольку кутикула после своего формирования расти не может, рост и изменение внешнего строения организма в ходе его онтогенеза обязательно сопровождаются линьками. Линька представляет собой полную одновременную смену кутикулы на всем теле. Старая кутикула сбрасывается после того, как под ней сформируется новая; происходит это следующим образом. Вначале происходит аполиз (apolysis) – гиподерма отслаивается от старой кутикулы, и пространство между гиподермой и кутикулой заполняется экзувиальной жидкостью. Внутренние слои старой кутикулы растворяются под действием ферментов, содержащихся в экзувиальной жидкости. На этом этапе возможен рост или дегенерация тех или иных участков гиподермы (см. ниже). Затем на поверхности гиподермы выделяется новая кутикула; в это время формируются все кутикулярные образования, свойственные будущему возрасту, в том числе артроподные щетинки и их производные [см. ниже, Euarthropoda (5)]. Затем происходит экдизис (ecdysis) – сбрасывание старой кутикулы.
Обычно старая кутикула сбрасывается целиком, так что животное вылезает из нее; сброшенная кутикулярная шкурка называется экзувий (латинское слово exuviae употребляется только во множественном числе). Экзувий включает весь наружный кутикулярный покров, а также кутикулярную выстилку передней и задней кишки и других внутренних структур эктодермального происхождения (трахеи, аподемы и др.).
У большинства насекомых при линьке старая кутикула трескается по медиальной продольной линии на дорсальной стороне тела [см. ниже, Hexapoda (14)]; у других членистоногих линии разлома кутикулы иные.
В процессе линьки может происходить как незначительное изменение размеров и строения, так и очень резкое увеличение или уменьшение тела или отдельных его частей, появление или исчезновение сегментов и придатков.
Увеличение наружного придатка происходит следующим образом. Когда гиподерма отслаивается от кутикулы, под старой кутикулой происходит рост и изменение формы гиподермы; поскольку кутикула при этом не увеличивается, растущая под ней гиподерма придатка утолщается и/или сминается в складки. Перед линькой выросший гиподермальный зачаток придатка помещается под старой кутикулой либо за счет того, что его гиподерма сжата, либо за счет того, что он смят в складки или петли (рис. 9.2.2, 9.10.3I, 10.1.1C). Такой гиподермальный зачаток бывает вдавлен в гиподерму туловища, образуя в ней карман; карман бывает базигенным (когда зачаток погружен в карман своим основанием – рис. 10.2.7A) или акрогенным (когда зачаток погружен в карман своей вершиной – рис. 10.4.7B). Форма сминания гиподермы под старой кутикулой может иметь таксономическое значение, поскольку иногда гомологичные органы, имеющие сходное дефинитивное строение, по-разному смяты перед линькой. У личинок круглошовных мух (Diptera-Cyclorapha) имеются имагинальные диски – гиподермальные зачатки будущих куколочных придатков, погруженные в своеобразные стебельчатые карманы; иногда термин «имагинальный диск» неудачно используют для любых других гиподермальных зачатков, а также в совершенно других значениях [см. ниже, Cyclorapha (1b)]. В некоторых случаях гиподермальный зачаток, погруженный в карман, существует не только в том возрасте, который непосредственно предшествует появлению наружного придатка, а закладывается раньше и переходит из возраста в возраст, не участвуя в очередной линьке и после линьки оставаясь внутри тела (рис. 10.1.3C–D); такой зачаток называется внутренним. После того, как гиподерма заканчивает свой рост, на ее поверхности формируется новая кутикула, повторяющая складки гиподермы; эта новая кутикула до линьки остается несклеротизованной, эластичной, в том числе в области склеритов [см. ниже, Euarthropoda (1)]. При выходе животного из старой кутикулы смятые участки гиподермы и новой кутикулы расправляются и приобретают форму, свойственную новому возрасту. Затем может происходить склеротизация новой кутикулы, приводящая к утрате ее эластичности.
Уменьшение придатка может происходить разными способами – антирост, обрыв без отходов или обрыв с отходами (см. Указатель признаков [5.10–12]). Антирост – это постепенное уменьшение и упрощение придатка, напоминающее эмбриональный рост и развитие в обратной последовательности; в результате антироста новый придаток занимает малую часть объема внутри старой кутикулы, но все его части происходят непосредственно из гомологичных частей старого придатка, и между новой гиподермой и старой кутикулой не остается отброшенных тканей (рис. 10.15.2A–C, 10.15.2D–F). Обрыв: старый придаток или его дистальная часть отрывается и оказывается в пространстве между новым придатком и старой кутикулой; обрыв без отходов: оторвавшаяся часть целиком растворяется и всасывается организмом, так что после линьки в сброшенном экзувии ничего не остается (рис. 10.4.7A–C, 10.4.7D–F, 10.4.7G–J); обрыв с отходами: при линьке остатки оторвавшейся части оказываются внутри сброшенного экзувия. В результате линьки с обрывом придаток либо оказывается лишенным дистальной части, либо имеет все те же части, что и до линьки; в последнем случае, в отличие от антироста, его части развиваются из негомологичных частей старого придатка. Линька с обрывом либо сопровождается нормальным аполизом, либо обрывающаяся часть не подвергается аполизу, т.е. вообще не участвует в линочном цикле и сбрасывается вместе с экзувием в неизмененном виде [в частности, так сбрасываются тергалии поденок при превращении личинки в субимаго – см. ниже, Ephemeroptera (12b)].
Иногда в ходе одной линьки орган последовательно претерпевает антирост и рост, либо обрыв и рост. Обычно такие процессы сопровождают линьку, приводящую к сильному изменению в строении органа; но в некоторых случаях сложные линочные процессы происходят при линьке, в результате которой строение органа не меняется [см. ниже, Gallinsecta (1)]. В некоторых случаях способ развития того или иного органа является очень консервативным: например, у всех Ephemeroptera при превращении личинки в субимаго дегенерация тергалий, антенн и парацерка (если он укорочен у крылатых стадий) происходит путем обрыва с отходами, а ротовые органы дегенерируют путем антироста [см. ниже, Ephemeroptera (1b)]. В других случаях гомологичные органы могут развиваться разными способами: так, например, происходит с ногами при превращении личинки в куколку у насекомых с полным превращением [см. ниже, Metabola (1Cc)].
У подавляющего большинства членистоногих рост покровов без линьки невозможен: если площадь поверхности тела увеличивается без линьки, то лишь за счет расправления складок или благодаря упругому растяжению. Известны лишь немногие исключения, когда мягкое туловище насекомого способно сильно увеличиваться за счет роста и размножения гиподермальных клеток и необратимого увеличения площади кутикулы (Зацаринина 1972).
Благодаря наличию линек, все постэмбриональное развитие членистоногого четко распадается на возрасты – промежутки между линьками. В пределах одного возраста размер и внешнее строение животного остаются неизменными, а все изменения (результаты роста, антироста или обрыва) становятся видны снаружи только после сбрасывания экзувия, то есть в следующем возрасте. Число возрастов бывает неопределенным (т.е. варьирует у особей одного вида), либо определенным; в некоторых случаях число возрастов является постоянным для крупного таксона (см. Указатель признаков [5.16]: табл. G).
Границей между возрастами принято считать момент сбрасывания старой кутикулы; каждому возрасту предшествует фаратная фаза (phasis pharata) этого возраста, когда новая кутикула, свойственная данному возрасту, уже развилась, но находится еще под кутикулой предшествующего возраста:
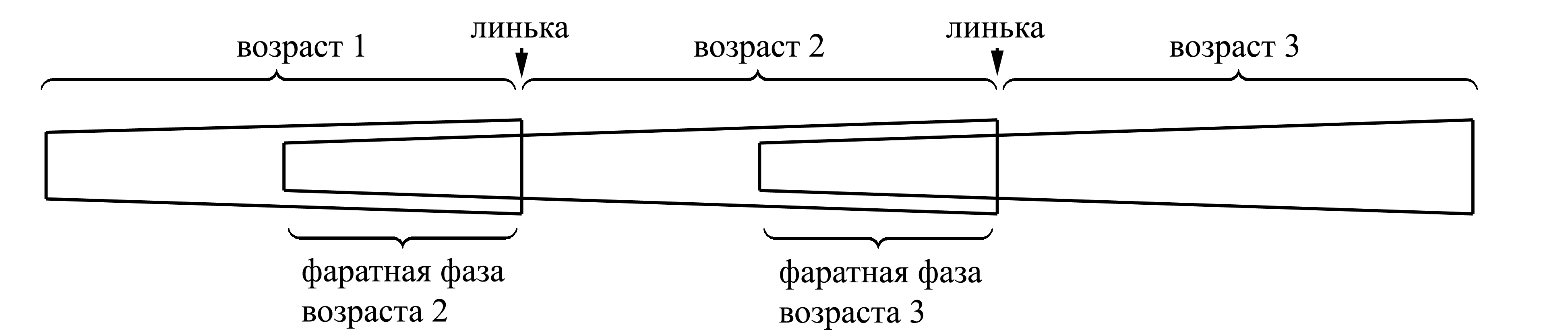
Принято считать возраста только у животного, покинувшего яйцевые оболочки; однако бывают и эмбриональные линьки, происходящие в яйце. Новорожденное животное [или личинка – см. ниже, Pterygota (4C)], выходя из яйца, может оставлять в яйце свои эмбриональные экзувии; но иногда из яйца выходит предличинка, или преларва (praelarva), покрытая эмбриональной кутикулой, и лишь затем она линяет на первый возраст.
У большинства членистоногих кутикула является не только покровом, но и скелетом [см. ниже, Euarthropoda (1)], благодаря чему в ходе некоторых линек возможно очень резкое изменение строения животного; это привело к появлению разных форм метаморфоза (metamorphosis), при котором постэмбриональное развитие разделяется на стадии. Стадией (stadium), или итером (iter), принято считать часть онтогенеза, охватывающую один или несколько возрастов, слабо различающихся по внешнему строению друг от друга и отделенных от другой стадии линькой, при которой внешнее строение сильно изменяется. В то время как число возрастов является абсолютно объективным, число стадий, насчитываемое разными исследователями, может различаться в зависимости от того, что тот или иной исследователь считает сильным изменением строения.
У некоторых членистоногих кутикула определенной стадии развития, отслоившаяся от гиподермы, не препятствует приобретению гиподермой новой формы и не вызывает образования предлиночных складок гиподермы; иногда сквозь отслоившуюся прозрачную кутикулу можно видеть, как гиподерма постепенно меняет форму, так что создается иллюзия постепенного превращения; однако и в этих случаях стадии развития разделены линьками, выражающимися в появлении новой кутикулы и сбрасывании старой кутикулы.
(3 c) Трахеи. В различных группах членистоногих, перешедших к наземному образу жизни, независимо возникла трахейная система, представляющая собой систему проходящих внутри тела трубок – трахей (tracheae), образованных эктодермальным эпителием и выстланных изнутри кутикулой. С наружными покровами трахейная система сообщается в устьях, которые предлагается далее называть трахеостии (tractheostia); часто трахеостия имеет вид отверстия и служит для газообмена между трахейной системой и окружающим воздухом, т.е. является дыхальцем; обычно трахеостия заметна на поверхности тела как пятнышко, или глазок – стигма (stigma). Дыхальце может располагаться как на мембранозном участке покрова, так и на склерите; если дыхальце окружено собственным кольцевидным склеритом, такой склерит называют перитремой (peritrema). В некоторых случаях трахеостия замкнута, не служит для газообмена и не заметна [см. ниже, Hexapoda (9) и Amyocerata (2)]. В трахеостиях эпителий трахеи переходит непосредственно в покровный эпителий, а кутикулярная выстилка трахеи переходит непосредственно в покровную кутикулу. В онтогенезе трахейная система закладывается как впячивания покровов на месте трахеостий, а при каждой линьке кутикулярная выстилка трахей сбрасывается вместе с покровной кутикулой, выходя наружу через трахеостию. У многих членистоногих трахеи разветвлены и анастомозируют внутри тела, так что трахеи, идущие от разных дыхалец, могут образовывать единую сеть или трахейные стволы, проходящие через все тело; в этом случае при линьке кутикулярная выстилка трахейной системы рвется на фрагменты, каждый из которых выходит через свою трахеостию.
Трахейная система имеется у онихофор, у различных паукообразных, у большинства многоножек и насекомых. У мухоловок (Chilopoda-Notostigmophora) и у некоторых мокриц [Isopoda-Terrestria Lattreille 1825 {nom.typ.: Oniscus/fg (incl. Tylos, Armadillo)}] имеются дыхательные органы, которые, так же как и трахеи, образованы впячиваниями покровов и выстланы тонкой кутикулой, но, в отличие от типичных трахей, не проникают глубоко в тело, а ветвятся в ограниченном пространстве – у мухоловок в небольшой области под туловищным тергитом [см. ниже, Notostigmophora (5)], а у мокриц внутри экзоподита абдоминальной конечности (Hoese 1982); эти дыхательные органы называют либо трахеями, либо легкими, либо «псевдотрахеями» (что не следует путать с псевдотрахеями в хоботке двукрылых).
Трахейная система возникла в разных таксонах членистоногих независимо, о чем свидетельствует разное положение трахеостий: поскольку трахейная система имеет эктодермальное происхождение и закладывается как впячивания покровов, положение трахеостий должно быть единым у животных, имеющих единое происхождение трахейной системы.
У онихофор трахеостии расположены беспорядочно [см. ниже, Onychophora (2)]. У разных паукообразных трахеостии открываются на разных сегментах опистосомы или просомы (см. ниже, Chelicerata: «Классификации» I: Caulogastra, Pseudoscorpii, Mycetophora, Phalangium/fg1 и Monomerosomata). У наземных ракообразных (Isopoda-Terrestria) трахеи, если имеются, расположены внутри определенных пар абдоминальных конечностей (плеоподов) и открываются на их поверхности. Среди Atelocerata, у симфил трахеостии открываются в мандибулярные аподемы; у диплопод и пауропод – в коксальные аподемы; у Chilopoda трахеостии расположены на туловище, но у Notostigmophora они непарные дорсальные, а у Pleurostigmophora – парные латеральные; у насекомых трахеостии латеральные и расположены по-разному на груди и на брюшке; среди насекомых у ногохвосток трахеи имеют независимое происхождение и открываются на шее [см. ниже, Atelocerata (6), Symphyla (2), Collifera (5), Notostigmophora (5), Pleurostigmophora (7), Hexapoda (9) и Collembola (4)].
Часто кутикулярная выстилка трахей образует спиральные утолщения – тенидии (taenidia), предохраняющее трахею от сплющивания при сохранении ее гибкости. Очень сходная форма тенидий имеется у ??многих Arachnida, у Chilopoda-Pleurostigmophora и у Hexapoda – т.е. у представителей Chelicerata и Mandibulata, независимо перешедших к наземному образу жизни и поэтому явно независимо приобретших трахейную систему.
(3 d) Кутикулярные образования кишечника. Кутикулярная выстилка эктодермальной передней кишки [стомодеума – см. Monostomata (2)] обычно равномерно тонкая и эластичная; но у некоторых членистоногих самая задняя часть стомодеума образует небольшой отдел – преджелудок, или провентрикулюс (proventriculus), который имеет особое кутикулярное вооружение – склеротизованные гребни, зубцы, шипы, аканты и др (рис. 10.4.3B, 10.8.1A). При линьке кутикулярная выстилка стомодеума выходит через ротовое отверстие. Кутикулярная выстилка эктодермальной задней кишки [проктодеума – см. Haemataria (1)] не имеет склеритов; при линьке она выходит через анальное отверстие. Эпителий средней кишки, будучи энтодермальным, кутикулу не вырабатывает; средняя кишка может быть выстлана изнутри тонкой пористой хитинсодержащей перитрофической мембраной, вырабатываемой эктодермальными клетками передней кишки.
(3 e) Хитин и кутикулы других животных. Способность вырабатывать хитин исходно присуща живым организмам [см. Eucaryota (5)]. Внеклеточная кутикула того или иного строения, помимо членистоногих, встречается в различных таксонах животных: у кольчатых червей (Annelida), моллюсков (Malacozoa), щупальцевых (Lophophorata) (см. Peritoneata: «Классификация»); у брюхоресничных (Gastrotricha), нематод (Nematoidea), головохоботных (Cephalorhyncha) (см. Haemataria: «Классификация») и др. В отличие от членистоногих, кутикула этих животных образована не хитином, а волокнами коллагена [см. Metazoa (1c)] или других белков. При этом организм этих животных способен вырабатывать хитин, который участвует в образовании тех или иных внешних структур (яйцевых оболочек, щетинок у кольчатых червей и щупальцевых, радулы у моллюсков); в некоторых случаях хитин присутствует и в составе кутикулы, где он может не быть обязательным компонентом: например, хитин отсутствует в кутикуле взрослых нематод, но содержится в небольших количествах во внутреннем слое кутикулы личинок; у приапулид (Priapulida, относящихся к Cephalorhyncha) хитин может появляться во внутреннем слое старой кутикулы и отсутствовать в молодой кутикуле.
В отличие от других животных, у Gnathopoda хитин составляет основу кутикулы. Кутикула членистоногих резко отличается от кутикулы других представителей Metameria – кольчатых червей.
У кольчатых червей (Annelida) имеется тонкая эластичная кутикула; в отличие от кутикулы членистоногих, она образована не хитином, а преимущественно коллагеном, который секретируется многочисленными микровиллями – тонкими отростками эпителиальных клеток; волокна коллагена откладываются между микровиллями, так что микровилли пронизывают кутикулу насквозь и их вершины выступают над поверхностью кутикулы. Поскольку микровилли откладывают коллаген и другие вещества кутикулы сразу по всей толще кутикулы, при росте червя его кутикула также растет в ширину, поэтому рост обычно происходит без линек; лишь у немногих представителей Annelida наблюдается линька, при которой сбрасывается вся кутикула или лишь ее часть. У кольчатых червей имеется хитин, но в отличие от членистоногих, хитин не входит в состав кутикулы, а содержится только в щетинках [см. ниже, Euarthropoda (5e)].
Помимо Gnathopoda, кутикула регулярно сбрасывается у Nematoidea и Cephalorhyncha; на основании этого некоторые авторы объединяют Gnathopoda, Nematoidea и Cephalorhyncha в таксон Ecdysozoa и считают наличие линяющей кутикулы не аутапоморфией Gnathopoda, а аутапоморфией Ecdysozoa (см. ниже, «Систематическое положение»).
(4) Конечности. Органами передвижения служат конечности – выросты тела, исходно расположенные по паре на каждом сегменте; предположительно, конечности Gnathopoda гомологичны параподиям Polychaeta [см. Metameria (3)].
В отличие от Gnathopoda, у Polychaeta органами передвижения в значительной мере служат подвижные аннелидные щетинки [см. ниже, Euarthropoda (5e)], расположенные на параподиях; в связи с этим параподии полихет иногда называют хетоподиями, а конечности Gnathopoda называют лобоподиями (lobopodia); иногда лобоподиями называют только нечленистые конечности Onychophora и Tardigrada, в отличие от артроподий настоящих членистоногих [см. ниже, Euarthropoda (3)].
Расположение конечностей строго по одной паре на сегменте очень консервативно; часто по расположению пар конечностей можно выявить гомологию сегментов в случаях их слияния или редукции. Исключение составляют диплосегментные многоножки, у которых один сегмент может нести две пары конечностей [см. ниже, Myriapoda (3)]. Другим исключением является наличие более чем по одной паре листовидных конечностей на туловищных сегментах у Notostraca (см. ниже, Gnathostraca: «Классификация»).
Локомоторные конечности обычно называют ногами (pedes); у разных Gnathopoda на тех или иных сегментах конечности бывают сильно видоизменены и приспособлены для иной функции, нежели локомоторная: конечности бывают ротовыми (различные пары челюстей – мандибул, максилл и др.), хватательными, сенсорными, дыхательными (жабры и др.), защитными, половыми и другими; иногда конечности тех или иных сегментов утрачены.
Локомоторные конечности обычно снабжены коготками (onychia, или ungues) – заостренными выростами со склеротизованной кутикулой. У Onychophora и Tardigrada конечности мягкие, а вершина каждой конечности исходно несет 2 коготка [см. ниже, Onychophora (1) и Tardigrada (3)]. У Euarthropoda конечности состоят из склеротизованных члеников, на вершине обычно с коготками и другими образованиями (см. Указатель признаков [2.58–63]).
В строении конечностей разных пар исходно проявляется эволюционная корреляция. В случаях глубокой дифференциации конечностей, их эволюционная корреляция может утрачиваться. В некоторых случаях эволюционная корреляция в строении конечностей разных пар возникает вторично [см. ниже, Hexapoda (1b)].
Помимо конечностей или их производных, сегмент может нести подвижные придатки иного происхождения; в частности, у насекомых, помимо пары конечностей, определенные сегменты несут крылья [см. ниже, Pterygota (4)] или тергалии [см. ниже, Pterygota (4D)]. Последний сегмент тела может нести каудалии, псевдокаудалии или другие придатки, не гомологичные конечностям [см. ниже, Hexapoda (8)]. Гомологию придатков не всегда легко определить, поэтому некоторые авторы ошибочно принимают за гомологи конечностей крылья, тергалии, каудалии и некоторые другие придатки.
Геологический возраст. Кембрий – ныне.
Систематическое положение Gnathopoda. Некоторые авторы возражают против объединения Gnathopoda и Annelida в таксон Metameria в составе Peritoneata; вместо этого они объединяют Gnathopoda с нецеломическими животными Nematoidea (нематоды) и Cephalorhyncha (головохоботные) в таксон Ecdysozoa Aguinaldo & al. 1997. Единственным общим признаком Ecdysozoa является наличие хитинсодержащей кутикулы, сбрасываемой при линьках [см. Gnathopoda (3)]. Кроме того, в качестве доказательства голофилии Ecdysozoa ссылаются на некие «молекулярные данные». Однако не указано ни одного молекулярного признака, общего для Gnathopoda, Nematoidea и Cephalorhyncha и отсутствующего у других животных, а под «молекулярными данными» понимают лишь результаты вычислений, основанных на принципе парсимонии (см. I.4.2.6 и I.4.2.7).
Наличие линяющей хитинсодержащей кутикулы, хотя и является признаком, общим для Gnathopoda, Nematoidea и Cephalorhyncha, но недостаточно для доказательства родства этих таксонов (Schmidt-Rhaesa 2007).
В отличие от кутикулы членистоногих, основу которой составляет хитин, кутикула нематод и головохоботных образована не хитиновыми волокнами, а волокнами белковой природы, сходными с коллагеном [см. Gnathopoda (3e)]. Нематоды и головохоботные, как и многие другие эукариоты, способны вырабатывать хитин [см. Eucaryota (5)], но присутствие хитина в их кутикуле не обязательно. Во внешней кутикуле взрослых нематод хитин отсутствует, а их кутикула считается «хитинсодержащей» только потому, что малые количества хитина обнаружены во внутреннем слое кутикулы молодых нематод; также кутикула приапулид считается «хитинсодержащей» только потому, что малые количества хитина появляются во внутреннем слое кутикулы на заключительной стадии ее созревания.
В разных таксонах животных независимо возникал линочный цикл, при котором сначала старая кутикула отслаивается от эпителия, затем эпителий выделяет новую кутикулу, а затем старая кутикула сбрасывается. О возможности независимого возникновения такого цикла свидетельствует то, что он встречается у некоторых кольчатых червей: линька туловищной кутикулы подробно описана для медицинской пиявки (Hirudo medicinalis Linnaeus 1758) и отмечена у некоторых полихет; у некоторых хищных полихет таким же способом в процессе роста животного заменяется не вся кутикула, а только участок глоточной кутикулы, несущий зубы.
Сходство между линькой хитиновой кутикулы членистоногих и линькой белковой кутикулы нематод не большее, чем их сходство с линькой белковой кутикулы пиявок. Судя по всему, в этих трех случаях линочный цикл возникал независимо.
В отличие от Metameria, у нематод и головохоботных метамерная сегментация отсутствует, а по общему плану строения (взаиморасположение нервной системы, кровеносной системы, мышц и других частей тела) они не имеют ничего общего с членистоногими [см. Metameria (1)–(5)].
Статус Gnathopoda. Помимо представления о голофилии Gnathopoda, высказывалась гипотеза о полифилетическом происхождении Gnathopoda, согласно которой разные группы Gnathopoda берут свое начало от разных Polychaeta; при этом полифилетическими таксонами считаются не только Gnathopoda, но и Euarthropoda и Mandibulata (см. Классификации Gnathopoda I) (Packard 1903, Manton 1964, 1977, Anderson 1973). В связи с этим некоторые авторы не признают таксон Gnathopoda или считают его не таксоном, а градой (т.е. лишь «уровнем организации»). В пользу голофилии Gnathopoda свидетельствуют апоморфии (см. выше), тогда как в пользу теории о полифилии Gnathopoda не приведено ни одного аргумента, кроме общих слов о том, что эти апоморфии могли возникать независимо несколько раз.
Классификации Gnathopoda
I. Деление Gnathopoda на Ceratophora и Acerata. Эта классификация обычно связывается с представлением о полифилии Gnathopoda (Manton 1964 и др.) (см. Статус Gnathopoda); при этом филогению Metameria представляют следующим образом:

Авторы, придерживающиеся такой гипотезы о филогении, считают таксоны Euarthropoda (т.е. членистоногих без Onychophora) и Mandibulata (т.е. Eucrustacea + Atelocerata) полифилетическими и игнорируют аутапоморфии, доказывающие голофилию каждого из них (см. ниже).
Таксон Acerata Lankester 1888 {syn.circ.: Cephalothoracica Goette 1902} в этой классификации является заведомо полифилетическим (если считать Gnathopoda полифилетической группой, как показано на схеме) или парафилетическим (если считать Gnathopoda голофилетической группой); в связи с этим его обычно не признают, рассматривая Eucrustacea и Chelicerophora как таксоны того же ранга, что и Ceratophora.
Таксон Ceratophora Lankester 1888 {synn.circ.: Eucephalica Goette 1902 = Uniramia Manton 1977} объединяет Onychophora и Atelocerata. Общими признаками этих таксонов являются лишь наземный образ жизни, наличие трахейной системы и габитуальное сходство Onychophora с многоножками (Myriapoda) и гусеницами (личинками Lepidoptera). В многочисленных работах С.М. Мэнтон (Manton 1977 и др.) и некоторых других авторов (Anderson 1973 др.) Ceratophora рассматривался как голофилетический таксон, однако ни одной аутапоморфии Ceratophora не описано. Разными авторами опубликованы результаты многих сравнительно-анатомических и сравнительно-эмбриологических исследований с целью выявить признаки родства Onychophora и Atelocerata, однако до сих пор не описано ни одного их общего признака, который при этом не был бы общим признаком всех Gnathopoda.
Наличие трахейной системы, которое характерно и для Atelocerata, и для Onychophora, не является их синапоморфией: трахейная система возникла в этих группах, а также у некоторых других членистоногих независимо, о чем свидетельствует разное положение дыхалец [см. Gnathopoda (3c)].
Высказывалась гипотеза о происхождении мандибул Atelocerata из ротовых крючьев Onychophora [о необоснованности этой гипотезы см. ниже, Mandibulata (1c)].
Аргументом в пользу независимого происхождения Chelicerata, Eucrustacea и Atelocerata от разных Polychaeta некоторые авторы считают разное количество ларвальных сегментов в этих таксонах [см. Metameria (1b)]. При этом почему-то предполагается, что только у Polychaeta число ларвальных сегментов может меняться в ходе эволюции, а у членистоногих оно должно оставаться неизменным и соответствовать числу ларвальных сегментов у предковой группы Polychaeta. Считается, что у ракообразных (Eucrustacea) в типичном случае позади антенн первых (т.е. в посторальном отделе тела) имеется 2 ларвальных сегмента, так как их личинка (науплиус) имеет помимо антенн I только две пары конечностей – антенны II и мандибулы [см. ниже, Eucrustacea (1) и рис. 4.7.1E]. Все насекомые и многоножки (Atelocerata), будучи наземными или вторичноводными животными, утратили первичную плавающую личинку. Поэтому о количестве ларвальных сегментов морских предков Atelocerata судят по тому, в какой последовательности развиваются сегменты современных Atelocerata в эмбриогенезе. У разных изученных в этом отношении Atelocerata эта последовательность различается. По мнению некоторых исследователей, наиболее типичным (и, как полагают, первичным) является наличие четырех «передних ларвальных» посторальных сегментов и еще трех «задних ларвальных» сегментов на конце тела (Иванов 1940). На самом деле вопрос о числе ларвальных сегментов как у Eucrustacea, так и у Atelocerata неясен, и разные авторы называют разное число в зависимости от того, что считают главным различием ларвальных и постларвальных сегментов (относительное время закладки или способ сегментации эктодермы или мезодермы, или другие признаки). В частности, согласно одной из интерпретаций, у всех Euarthropoda, включая Eucrustacea, имеется одинаковое число – 4 ларвальных сегмента, хотя не всегда все они закладываются одновременно (Snodgrass 1938 и др.).
На самом же деле различия в числе закладывающихся одновременно («ларвальных») сегментов можно объяснить, не прибегая к гипотезе о полифилетическом происхождении членистоногих. Очевидно, что изменение последовательности закладки сегментов может происходить не только в эволюции Polychaeta, но и в эволюции Atelocerata; в частности, у некоторых насекомых все сегменты закладываются одновременно, т.е. ведут себя как «ларвальные». Предполагают, что увеличение числа одновременно закладывающихся сегментов – общая тенденция, связанная с эмбрионизацией развития, но в ряде случаев возможен и обратный процесс (Захваткин 1975). Поскольку «передними ларвальными» называют сегменты головы, а «задними ларвальными» – генитальные сегменты, отличие этих сегментов от «постларвальных» у эмбриона может быть связано не с памятью о древних предках, а со специализацией этих сегментов у взрослого животного. Кроме того, изменение числа ларвальных сегментов (т.е. сегментов, имеющихся у первичной плавающей личинки) могло происходить и у морских предков Atelocerata, которые, очевидно, такую личинку имели.
Эмбриональное развитие членистоногих настолько разнообразно, что высказывались даже соображения о том, что Eucrustacea имеют происхождение, независимое не только от Atelocerata, но даже от Polychaeta (Anderson 1973). Однако большое разнообразие способов дробления, гаструляции, образования мезодермы и сегментации иногда наблюдается даже в пределах небольшого таксона, так что оценить филогенетическое значение этих признаков трудно. Некоторые авторы ошибочно полагают, что признаки, относящиеся к наиболее ранним стадиям онтогенеза, должны прояснять филогенетические связи наиболее крупных таксонов; эта точка зрения связана со старинным «биогенетическим законом», не согласующимся с современными представлениями о механизмах наследственности и эволюции (см. I.4.2.1).
II. Деление Gnathopoda на Onychophora и Euarthropoda. Здесь мы придерживаемся этой классификации, основанной на представлении о голофилии Gnathopoda и голофилии Euarthropoda (см. ниже).
Некоторые авторы объединяют Euarthropoda и Tardigrada в таксон Arthropoda Siebold 1848 {synn.circ.: Arthropodoidea De Haro 1999 = Tactopoda Budd 2001 = Tritocerebra Grimaldi & Engel 2005}. Возможной аутапоморфией Arthropoda является наличие тергитов и стернитов и утрата кольцевых мышц [см. ниже, Tardigrada (4) и Euarthropoda (2)].
Иногда Onychophora и Tardigrada объединяют в плезиоморфон Pararthropoda Vandel 1949, который характеризуется лишь отсутствием характерных для Euarthropoda признаков.
В качестве таксона, непосредственно подчиненного Gnathopoda, иногда рассматривают также пятиусток – Acanthotheca Diesing 1835 {syn.circ: Maeracara Haeckel 1889; nom.typ.: Linguatula/fg [f:1859; g:1789]}. Строение пятиусток сильно модифицировано в связи с приспособлением к паразитизму на позвоночных, поэтому их систематическое положение неясно; некоторые авторы относят их к Eucrustacea-Copepodoidea.
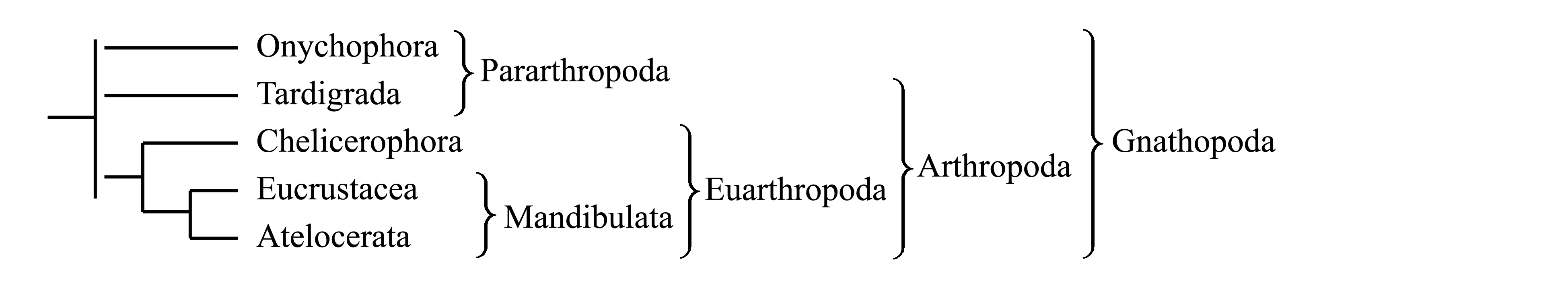
Здесь мы принимаем в составе Gnathopoda три подчиненных таксона – Onychophora, Tardigrada (стр. 000) и Euarthropoda (стр. 000).
Gnathopoda, или Araneus/fg6
![]() 1. Onychophora, или Peripatus/fg1
1. Onychophora, или Peripatus/fg1
![]() 2. Tardigrada, или Macrobiotus/fg1
2. Tardigrada, или Macrobiotus/fg1
![]() 3. Euarthropoda, или Araneus/fg7
3. Euarthropoda, или Araneus/fg7