КЛАДОЭНДЕЗИС НАСЕКОМЫХ

![]()
|
КЛАДОЭНДЕЗИС НАСЕКОМЫХ |
|||||||
|
|
|
fg | |||||
IV-1.3. Euarthropoda Lankester 1904 – настоящие членистоногие
(рис. 4.4.1–10.16.2)
|
Названия, однозначно соответствующие по циркумскрипции: — Condylopodes Bronn 1850 (non Condylopa Latreille 1825); — Arthropodes Bronn 1850 (non Arthropoda Siebold 1848); — Euarthropoda Lankester 1904. Названия, неоднозначно соответствующие по циркумскрипции (соответствующие также Gnathopoda): — Condylopoda Newman 1835, s.str.; — Endomyaria Spinola 1850, s.str. Иерархическое название: Araneus/fg7 [fg:(1757)1758] (sine Peripatus, Macrobiotus; incl. Scarabaeus). |
|
Ранг. В XIX–XX веках – обычно тип или подтип; в XVIII веке – обычно класс.
Название. Иногда этому таксону приписывают название «Arthropoda». В действительности название Arthropoda Siebold 1848 исходно было дано таксону, который, помимо настоящих членистоногих, включал тихоходок в качестве подотряда Tardigrada в составе класса Arachnida (Siebold 1848). Таким образом, название Arthropoda по своей первоначальной циркумскрипции соответствует таксону, объединяющему Euarthropoda и Tardigrada (см. Gnathopoda: «Классификации» II).
Аутапоморфии Euarthropoda
(1) Наружный скелет. Хитинсодержащая кутикула, покрывающая тело [см. Gnathopoda (3)], дифференцирована на склериты (от «scler...») (участки жесткой, т.е. склеротизованной, кутикулы) и мембраны (membranae) (эластичные участки), благодаря чему кутикула образует наружный скелет. Многие мышцы крепятся к склеритам таким образом, что при их сокращении происходит сгибание покрова по мембранам; расположенные тем или иным образом склериты, мембраны и мышцы функционируют как рычажные механизмы.
Наружный скелет и связанная ним мускулатура имеют свои особенности на туловище [см. (2)] и конечностях [см. (3)]. Туловище, конечности и антенны [см. (6)] имеют членистое (сегментированное) строение благодаря тому, что их кутикула дифференцирована на склеротизованные членики (сегменты) и эластичные сочленовные мембраны. Многие членики являются эволюционно консервативными, так что их гомология прослеживается в пределах крупных таксонов. В ходе эволюции происходило как разделение члеников, так и их слияние (некоторые авторы необоснованно пытаются представить всю эволюцию членистоногих только как слияние члеников). В некоторых случаях членики сливаются так, что граница между ними исчезает, и о происхождении членика из двух слившихся можно судить только на основании сравнения разных животных. В других случаях слившиеся членики, образуя монолитный склерит, в то же время сохраняют свою исходную форму и видимую границу между члениками; такие членики можно называть синсклеротичными.
Склериты туловища [см. (2a)], а также склериты конечностей [см. (3)] могут иметь разнообразные отростки, называемые апофизы (apophyses). Наружные апофизы, как и другие участки тела, имеют гиподерму внутри и кутикулу снаружи. Внутренние апофизы, называемые также аподемы (apodemata), представляют собой впячивания покровов, так что в них гиподерма находится по периферии, а кутикула выстилает ее изнутри; снаружи вход в аподему может иметь вид ямки или устья канала. Разнообразные аподемы, независимо возникшие у разных членистоногих, представляют собой эндоскелет, и часто служат рычагами, к которым крепятся мышцы. Иногда аподемы называют «хитиновыми сухожилиями». Следует отметить, что помимо эктодермальных эндоскелетных элементов (аподем), у членистоногих бывают в той или иной мере развиты настоящие сухожилия – мезодермальные эндоскелетные элементы, также служащие для прикрепления мышц. Вероятно, в ходе эволюции некоторых групп членистоногих отдельные мезодермальные сухожилия пространственно и функционально замещались аподемами, врастающими снаружи.
В некоторых группах членистоногих наличие сбрасываемого скелета привело к появлению метаморфоза [см. Gnathopoda (3b) и ниже, Pterygota (4C)].
(2) Скелет и мускулатура туловища. На туловище наружный скелет [см. (1)] включает прежде всего тергиты [см. (2a)], в связи с чем утрачены кольцевые туловищные мышцы [см. (2b)]. По наличию туловищных склеритов и утрате кольцевых мышц Euarthropoda отличаются от Onychophora; то же у тихоходок [ср.: Tardigrada (4)]; возможно, синапоморфия.
(2 a) Туловищные склериты. Исходно у каждого сегмента [см. Metameria (1)] дорсальная сторона – тергум (tergum), или нотум (notum) – несет склерит, называемый тергит. Вентральная сторона сегмента – стернум (sternum) – может нести склерит, называемый стернит. Латеральная стенка сегмента, расположенная между тергитом и стернитом – плейрон (pleuron), может быть мембранозной или образует склерит, называемый плейритом.
Границы склеротизации тергитов и стернитов изменчивы; иногда тергит и стернит сегмента слиты в цельное кольцо, иногда это кольцо разделено на вторичные склериты.
Часто продольные дорсальные мышцы [см. (2b)] крепятся к переднему краю тергита, а задний край тергита, свободный от мышц, налегает на тергит следующего сегмента. В связи с этим передний край тергита может нести антекосту (antecosta) – поперечное утолщение на внутренней стороне, служащее местом прикрепления продольных мышц; иногда антекоста находится отступя от переднего края тергита; в этим случае область тергита между его передним краем и антекостой называют акротергит (от «acro-» и «tergum»). Подобное строение встречается и у стернитов, где может быть развита антекоста, отделяющая впереди себя акростернит.
Иногда тергиты тех или иных сегментов шире самих сегментов, так что по бокам тела выступают плоские лопасти, называемые параноты (paranota); некоторые авторы считают параноты разных членистоногих гомологичными друг другу и связывают с ними гипотезы о происхождении крыльев насекомых [см. ниже, Pterygota (4E)]; однако вероятнее всего, что параноты многократно возникали и исчезали в ходе эволюции членистоногих.
(2 b) Туловищная мускулатура. В связи с дифференциацией кутикулы на склериты и мембраны кожно-мускульный мешок [см. Metameria (5)], распался на отдельные пучки мышц, приводящие в движение склериты. В частности, всегда отсутствуют кольцевые туловищные мышцы: их дорсальные и вентральные участки утрачены, а латеральные участки преобразованы в дорсовентральные мышцы, соединяющие тергит со стернитом. Так что в медиальной части тела непосредственно под покровами лежат продольные дорсальные и продольные вентральные мышцы, а в латеральной части – дорсовентральные мышцы (рис. 7.2.5E).
У некоторых Euarthropoda склериты вторично утрачены. В частности, многие личинки насекомых с полным превращением (Metabola) частично или полностью утратили туловищные тергиты и стерниты; некоторые из них, кроме того, утратили конечности [см. (3)] и двигаются как черви. Но и в этом случае кольцевые мышцы не восстанавливаются, и все движения туловища осуществляется за счет продольных, дорсовентральных и производных от них косых мышц.
(3) Членистые конечности. Парные придатки сегментов [см. Gnathopoda (4) и Metameria (3)] преобразованы в членистые конечности, называемые артроподии (arthropodia). Благодаря тому, что кутикула дифференцирована на склериты и мембраны [см. (1)], артроподия состоит из последовательно соединенных члеников (articuli), называемых также сегментами (segmenta), или подомерами (podomeri, или podomera): кутикула каждого членика обычно имеет вид цельного цилиндрического склерита, а следующие друг за другом членики разделены сочленением с мембранозной кутикулой. Каждый последующий членик, как правило, приводится в движение мышцами, находящимися в предыдущем членике. Один или несколько наиболее проксимальных члеников конечности двигаются мышцами, идущими из туловища – от тергита [см. (2a)], стернита, плейрита, мезодермального эндоскелета и/или тех или иных туловищных аподем. Число члеников конечности может быть различным [см. (3a)]; при этом не вполне ясно, какое число является исходным для членистоногих. В ходе эволюции отдельных групп членистоногих число члеников может уменьшаться или увеличиваться. Поскольку способность к регенерации конечностей оказала большое влияние на эволюцию наиболее крупной группы насекомых, здесь следует назвать общие особенности регенерации конечностей у членистоногих [см. (3b)].
(3 a) Сегментарный состав конечности. Предполагают, что исходно конечность совмещала жевательную и локомоторную функции (иногда такую конечность называют гнатопод); такая жевательно-локомоторная конечность имеет первый (т.е. наиболее проксимальный) членик, называемый коксоподит (от «coxa» и «pod-») и дистальную членистую часть, называемую телоподит (от «telo-»). Коксоподит исходно снабжен направленным медиально жевательным выступом – гнатобазой (gnathobasis), а телоподит часто используется для локомоции. Во многих случаях строение и функции конечности вторично изменены (см. «Классификации»). Иногда проксимальнее коксоподита имеются структуры, которые могут быть либо результатом вторичного расчленения, либо остатком особого членика – субкоксы (subcoxa) [см. ниже, Eucrustacea (2), Hexapoda (5b)].
У вымерших Trilobita и других Trilobitomorpha все конечности или часть из них имеют коксоподит с гнатобазой и членистый телоподит [см. ниже, Trilobitomorpha (1) и (2)]. У хелицеровых и морских пауков первая пара конечностей преобразована в 3-члениковые клешни или подверглась дальнейшей модификации [см. ниже, Chelicerophora (2)]; конечности следующих пар исходно 8-члениковые, иногда с уменьшенным или увеличенным числом члеников [см. ниже, Chelicerophora (4)]. У ракообразных, многоножек и насекомых конечности II пары преобразованы в мандибулы [см. ниже, Mandibulata (1)]. У ракообразных конечности исходно двуветвистые, с двучлениковой неразветвленной частью и двумя членистыми ветвями – экзоподитом и эндоподитом [см. ниже, Eucrustacea (2)]. У насекомых и многоножек конечности I пары утрачены, а конечности II пары преобразованы в одночлениковые мандибулы [см. ниже, Atelocerata (1) и (4)]; конечности III пары также преобразованы в ротовые придатки того или иного строения [см. ниже, Mandibulata (2), Symphyla (3), Collifera (1), Chilopoda (2), Hexapoda (2)]. У симфил конечности IV пары преобразованы в непарный нерасчлененный хилярий (нижнюю губу), а конечности V–XVI пар представляют собой 6-члениковые ходильные ноги [см. ниже, Symphyla (4) и (11)]. У диплопод и пауропод конечности IV пары утрачены [см. ниже, Collifera (2)], а ходильные конечности V и следующих пар у диплопод 8-члениковые, у пауропод 6-члениковые [см. ниже, Diplopoda (5)] и Pauropoda (6)]. У хилопод конечности, начиная с IV пары, имеют единый план расчленения и исходно 7-члениковые [см. ниже, Chilopoda (3), (4) и (7)]. У насекомых конечности IV пары преобразованы в непарную нижнюю губу усложненного строения, конечности V–VII пар представляют собой исходно 6-члениковые ходильные ноги, а конечности следующих пар утрачены или преобразованы в нерасчлененные придатки [см. ниже, Hexapoda (3), (5) и (7)]. Гомология между члениками конечностей у Hexapoda, Chilopoda, Pauropoda, Diplopoda, Symphyla, различных Eucrustacea и различных Chelicerophora в значительной мере неясна (рис. 4.8).
(3 b) Регенерация конечностей. У многих членистоногих конечности способны к регенерации (Maruzzo et al. 2005). В этом случае, если в результате травмы часть конечности утрачена, из оставшихся члеников этой конечности может развиться полноценная конечность с полным набором члеников. Такая регенерация возможна только при условии, что после травмы происходит одна или несколько линек, т.к. без линек внешние морфологические преобразования невозможны [см. Gnathopoda (3b)]. Ткани одного или нескольких проксимальных члеников, сохранившихся после утраты дистальной части конечности, сначала утрачивают свое исходное расчленение и формируют новое расчленение, так что образуют маленькую конечность с полным набором члеников; затем эта конечность растет, достигая размера нормальной нетравмированной конечности. Иногда весь процесс происходит под старой кутикулой до очередной линьки, так что в ходе одной линьки из культи сразу получается полноценная конечность. Иногда полная регенерация происходит за несколько линек, так что в некоторых возрастах на месте культи имеется маленькая конечность с полным набором члеников. Таким образом, при регенерации все членики конечности, включая дистальные, развиваются из тканей, принадлежавших одному или нескольким проксимальным членикам, т.е. не все филогенетически гомологичные членики оказываются онтогенетически гомологичными.
При нормальных линьках нетравмированных конечностей, как правило, каждый членик следующего возраста образуется из гомологичного членика предыдущего возраста, т.е. все филогенетически гомологичные членики являются онтогенетически гомологичными. Если в ходе линьки конечность увеличивается в размере, то обычно после аполиза каждый ее членик растет, сминаясь в пределах старой кутикулы этого же членика; благодаря этому находящиеся в нем мышцы сохраняют прикрепление к своим участкам кутикулы или кутикулярным аподемам [см. (1)], так что на протяжении всего этого процесса конечность сохраняет способность двигаться с помощью мышц и функционировать (рис. 7.2.1A, 9.12.3B). В случаях, когда из-за роста тканей конечности какое-либо сочленение сдвигается по отношению к старой кутикуле, такое сочленение утрачивает подвижность и остается неподвижным до сбрасывания старой кутикулы.
В некоторых таксонах членистоногих линочный процесс определенных пар конечностей преобразован так, что после аполиза ткани конечности сначала частично лизируются и утрачивают исходное расчленение, а затем заново приобретают расчленение и растут. В этом случае в период между аполизом и экдизисом конечность оказывается неподвижной. В результате такой линьки конечность приобретает членики, филогенетически гомологичные членикам предыдущего возраста, но онтогенетически не гомологичные им. Такой линочный процесс, сопровождаемый частичным лизисом, дедифференциацией и новой диффенциацией члеников конечности, возникал независимо несколько раз; в частности, он выявлен у личинок кокцид [см. ниже, Gallinsecta (1)] и при личиночоно-куколочной линьке насекомых с полным превращением [см. ниже, Metabola (1Cc)]. Вероятно, он обеспечивается теми же механизмами, что и процесс регенерации, при котором происходит заново дифференцируются членики конечности.
Высказывалось утверждение, будто при некоторых линьках, в частности при личиночно-куколочной линьке у Metabola, конечности могут возникать заново из каких-то зачатков, не относящихся к тканям конечности предыдущего возраста. Эти утверждения ошибочны, т.к. все конечности Euarthropoda (как, вероятно, и Metameria в целом), закладываются в раннем эмбриогенезе и повторно не образуются. По всей видимости, как в ходе нормального онтогенеза, так и в ходе регенерации после травм, все филогенетически гомологичные конечности оказываются онтогенетически гомологичными; в отличие от конечности в целом, филогенетически гомологичные членики конечности могут быть или не быть онтогенетически гомологичными.
(4) Зрительный аппарат. Имеется два зрительных аппарата; оба расположены на переднем отделе тела (т.е. на голове или головогруди) и иннервируются от протоцеребрума надглоточного ганглия [см. (6) и Metameria (2)], но от разных его отделов (рис. 6.6.2C): один зрительный аппарат исходно представляет собой пару фасеточных глаз и является уникальным новообразованием членистоногих; второй зрительный аппарат состоит из одной или нескольких пар глазков, предположительно гомологичных глазам онихофор и других животных.
Глаза (или латеральные глаза) (oculi) иннервируются от расположенных латерально оптических долей надглоточного ганглия. Обычно они имеют фасеточное строение, и в этом случае называются фасеточными, или сложными глазами. Каждый фасеточный глаз состоит из некоторого числа собранных вместе простых глаз – омматидиев (ommatidia); кутикулярные линзы омматидиев, сомкнутые вместе и образующие поверхность сложного глаза, называются фасетками (от французского facette). Глаза с таким фасеточным строением имеются во всех основных группах Euarthropoda если не у всех, то хотя бы у наиболее примитивных представителей: они есть у многих Trilobitomorpha, Eucrustacea, Amyocerata; среди Chelicerata фасеточные глаза имеются у Xiphosura, у палеозойских Gigantostraca и у некоторых палеозойских Toxicura. Все это свидетельствует о том, что фасеточные глаза были у общего предка членистоногих. У Arachnida, Myriapoda, Entognatha, личинок Metabola и некоторых других членистоногих вместо фасеточных глаз имеются нетипичные сложные (агрегированные) глаза или отдельные однолинзовые глаза – стеммы (stemmata), являющиеся результатом редукции фасеточных глаз. По набору зрительных клеток однолинзовый глаз может соответствовать одному омматидию, либо многим омматидиям сложного глаза, либо несопоставим со сложным глазом. В ходе постэмбрионального развития сложный глаз обычно увеличивается благодаря добавлению новых фасеток с каждым возрастом; новые фасетки добавляются по краям или с одного края; такое развитие глаз, вероятно, является исходным. В некоторых случаях число омматидиев с возрастом не меняется или скачкообразно увеличивается при определенной линьке; в некоторых случаях при определенной линьке весь глаз заменяется на новый (см. Указатель признаков [1.20]).
Глазки (ocelli) иннервируются от особого глазкового центра надглоточного ганглия, расположенного медиальнее зрительных долей. У Euarthropoda могут присутствовать две пары медиальных (фронтальных) глазков, пара вентральных глазков и пара дорсальных глазков (Paulus 1979). Одновременно бывают развиты до двух пар глазков. В отличие от глаз, глазки никогда не имеют фасеточного строения. Число пар глазков может быть сокращено; иногда пара глазков слита в непарный глазок. Глазки имеются у многих Trilobitomorpha, Chelicerata, Eucrustacea и у многих насекомых [см. ниже, Amyocerata (4)]; это говорит о том, что они были и у общего предка членистоногих.
Все фоторецепторные клетки глаз и глазков рабдомерные [см. Monostomata (3b)]. Как и в глазах некоторых аннелид и моллюсков, в глазах и глазках членистоногих фоторецепторные клетки обычно образуют один слой (называемый сетчаткой или ретинулой, по аналогии с глазами позвоночных) и вытянуты перпендикулярно плоскости этого слоя; рабдомеры занимают длинные боковые стенки фоторецепторных клеток таким образом, что образующие их микроворсинки направлены перпендикулярно оси клетки; во многих случаях соседние клетки плотно прилегают друг к другу своими рабдомерами, так что их рабдомеры образуют рабдом. В остальном как глаза, так и глазки различаются у разных членистоногих: они могут иметь различное число и расположение фоторецепторных клеток и разные конфигурации рабдомов; если глаза простые, они могут по строению не отличаться от глазков, отличаясь только иннервацией.
В онтогенезе многих членистоногих глаза и глазки появляются одновременно. У многих ракообразных глазки появляются раньше глаз; глазки ракообразных обычно называют науплиальными глазами, поскольку они присутствуют начиная со стадии науплиуса [см. ниже, Eucrustacea (1)]. У многих насекомых, наоборот, глаза появляются раньше глазков [см. ниже, Amyocerata (4)].
У многих членистоногих зрительный аппарат в той или иной степени редуцирован: могут быть утрачены либо глазки, либо фасеточные глаза, либо обе эти глазные системы (см. Указатель признаков [1.19–20]). При этом у Chelicerophora редукция обычно начинается с фасеточных глаз, так что у многих представителей глазки сохраняются, а фасеточные глаза либо редуцированы до нескольких или одной пары простых глаз, либо утрачены [см. ниже, Pantopoda (7), Chelicerata (2) и табл. IV.3]. У Atelocerata, наоборот, редукция всегда начинается с глазков: у всех Myriapoda и Hexapoda-Entognatha глазки утрачены, а глаза либо имеют нетипичное строение, либо тоже утрачены; у всех Hexapoda-Amyocerata, имеющих глазки, имеются развитые фасеточные глаза, тогда как у представителей с нефасеточными глазами или без глаз глазки всегда утрачены [см. ниже, Amyocerata (4)].
У некоторых других Metameria (Polychaeta, Onychophora, Tardigrada) бывает развита одна или несколько пар зрительных органов; возможно, эти органы, обычно называемые «глазами», гомологичны глазкам (ocelli) членистоногих, тогда как глаза (oculi) членистоногих являются новообразованиями и не гомологичны глазам каких-либо других животных (Mayer 2006). В глазах Onychophora и Polychaeta функцию светопреломления выполняет внеклеточное вещество, находящееся между покровом и слоем зрительных клеток. В глазках некоторых насекомых между покровом и слоем зрительных клеток также имеется внеклеточная полость; у других насекомых такой полости нет, и функцию светопреломления выполняет либо линзовидное утолщение покровной кутикулы, либо стекловидное тело, образованное эпителиальными клетками (Hallberg & Hagberg 1986).
Настоящие сложные глаза не встречаются ни у каких животных, кроме Euarthropoda. Глаза, напоминающие сложные, имеются на жаберных щупальцах некоторых сидячих полихет (Polychaeta-Sedentaria) и по краю мантии некоторых двустворчатых моллюсков (Bivalvia) (но не на голове этих животных); подобно сложному глазу, каждый такой глаз состоит из многих (50–150) зрительных элементов; в отличие от настоящего сложного глаза, каждый зрительный элемент состоит не из комплекса рабдомерных фоторецепторных клеток, а из единственной фоторецепторной клетки особого сложного строения: эта клетка имеет полость, в которую вдается множество фоторецепторных ресничек, расширенных в виде пластин (Nilsson 1994).
(5) Щетинки. На покровах имеются особые образования – артроподные щетинки (рис. 4.4.1A–C). Исходно артроподная щетинка представляет собой орган чувства – сенсиллу (sensillum), благодаря которому эктодермальная сенсорная клетка [см. Monostomata (3b)], будучи полностью покрытой кутикулой [см. Gnathopoda (3)], сохраняет способность воспринимать сигналы из внешней среды. Кутикула сенсиллы секретируется тремя концентрическими оболочечными клетками, окружающими сенсорную клетку: наиболее наружная из оболочечных клеток (тормоген) секретирует кольцевидную сочленовную мембрану вокруг основания щетинки, более внутренняя оболочечная клетка (трихоген) секретирует саму щетинку, а самая внутренняя оболочечная клетка (текоген) секретирует кутикулярный футляр, окружающий сенсорную ресничку (Harris 1977, Zacharuk 1985, Keil 1997). Обычно артроподную щетинку называют просто щетинка, или сета (латинское – saeta или seta), или хета (греческое – chaeta, χαιτη); иногда ее называют макротрихия (macrotrichion), в отличии от микротрихии [см. (5d)]. Артроподные щетинки имеют общие особенности строения [см. (5a)] и онтогенеза [см. (5b)]; при этом они расположены на самых разных участках покровов, имеют самую разнообразную форму и размеры, и выполняют самые разнообразные функции – различные виды механорецепции, хеморецепции, секреции и различные механические функции [см. (5c)]. Артроподные щетинки не следует смешивать с другими, внешне сходными образованиями на покровах тех же животных [см. (5d)] и с внешне сходными образованиями, имеющимися у некоторых других животных [см. (5e)].
(5 a) Строение артроподной щетинки (рис. 4.4.1A–C). Сенсорные и оболочечные клетки, образующие щетинку, лежат в общем слое гиподермы, так что каждая из них своим дистальным концом примыкает к кутикуле, покрывающей гиподерму.
Сенсорная клетка [сенсорный нейрон – см. Monostomata (3b)] имеет один аксон, отходящий от проксимального конца перикариона и тянущийся к центральной нервной системе [см. Metameria (2)], и один дендрит, отходящий от дистального конца перикариона и направленный к кутикуле; от вершины дендрита отходит неподвижная сенсорная ресничка (обычно одна). Ресничка имеет узкое основание с правильно расположенными девятью двойными микротрубочками [см. Eucaryota (2b)] и утолщенную дистальную сенсорную часть с неправильно расположенными микротрубочками. Исходно ресничка выполняет механорецепторную функцию: в ее дистальной части находится рецепторный участок, в котором расположено трубчатое тело, состоящее из множества параллельных микротрубочек, скрепленных друг с другом и с клеточной мембраной. Рецепторный участок реснички касается кутикулы щетинки и реагирует на изменения ее положения.
Оболочечные клетки последовательно намотаны на сенсорную клетку (или сразу на весь комплекс сенсорных клеток, если их несколько); каждая оболочечная клетка образует один полный виток, так что ее противоположные края плотно смыкаются. Между самой внутренней оболочечной клеткой (текогенной) и дендритом имеется внеклеточная полость – цилиарный синус; вокруг него в оболочечной клетке бывает развит сколопаль (scolopale), или сколопс (scolops) – плотное цитоплазматическое образование, укрепляющее стенки цилиарного синуса.
Особенностью артроподной щетинки является то, что ее кутикула [см. Gnathopoda (3)] формируется выделениями не менее чем трех специализированных оболочечных клеток – текогенной, трихогенной и тормогенной.
(A) Текогенная клетка (cella thecogena) (или текоген) – самая внутренняя оболочечная клетка, окружающая сенсорную клетку и образующая цилиарный синус. Текогенная клетка формирует кутикулу футляра, в котором заключена сенсорная ресничка. Этот трубчатый футляр (theca) состоит из очень тонкой эластичной кутикулы и своим дистальным концом соединен с кутикулой щетинки. Текогенная клетка, секретирующая кутикулу футляра, находится кнаружи от него, так что внутренний слой кутикулы футляра, обращенный к ресничке, соответствует наружному слою кутикулы насекомого.
(B) Трихогенная клетка (cella trichogena) (или трихоген) окружает текогенную клетку. Секретируемая ею кутикула образует собственно щетинку, т.е. склеротизованный вырост кутикулы.
(C) Тормогенная клетка (cella tormogena) (или тормоген) окружает трихогенную клетку и является наиболее наружной оболочечной клеткой. Она секретирует кольцевидную мембранозную область кутикулы, окружающую основание щетинки и обеспечивающую подвижность щетинки.
Проксимальнее оболочечных клеток обычно имеется одна или несколько глиальных клеток; они, как и оболочечные клетки, окружают нейрон, но, в отличие от оболочечных клеток, не выходит к поверхности и не секретирует кутикулу. Глиальные клетки окружают аксон и отчасти перикарион сенсорной клетки; они аналогичны клеткам неврилеммы (Шванновским клеткам), образующим миелиновую оболочку у позвоночных (Vertebrata) и некоторых других животных.
В некоторых сенсиллах число сенсорных и/или оболочечных клеток больше, чем названо выше. Сенсорных клеток может быть больше одной: обычно механорецепторная щетинка имеет одну или две сенсорные (механорецепторные) клетки, тогда как хеморецепторная щетинка – одну или две механорецепторные и много хеморецепторных клеток. Обычно каждая сенсорная клетка имеет лишь одну сенсорную ресничку, но иногда две. Кутикула щетинки обычно образуется одной трихогенной и одной тормогенной клеткой, но бывает большее число трихогенных и/или тормогенных клеток.
Строение кутикулярной части щетинки бывает разнообразным. Иногда внутри центрального канала имеется дополнительный кутикулярный рукав, внутри которого проходят реснички вместе с их кутикулярными футлярами. Наружный кутикулярный покров щетинки бывает полностью замкнутым, либо имеется линочная пора [см. (5b)], либо имеются дополнительные поры (наличие открытых пор характерно для хеморецепторных щетинок). Благодаря функционированию кольцевой тормогенной клетки щетинка обычно четко отчленена от остальной кутикулы, сидит в особом гнезде и имеет пассивную подвижность; но иногда щетинка неподвижно слита с остальной кутикулой.
Мышцы к щетинке никогда не подходят, так что подвижность щетинки, обеспечиваемая кольцевидной мембраной вокруг ее основания, может быть только пассивной.
(5 b) Развитие артроподной щетинки. Сенсорный нейрон и его оболочечные клетки образуются из одной предшествующей эктодермальной клетки в следующей последовательности:

При формировании кутикулы щетинки трихогенная клетка образует дистальный отросток, который секретирует кутикулу кнаружи от себя (рис. 4.4.1C). Когда процесс формирования кутикулы завершается, этот отросток трихогенной клетки дегенерирует, так что внутри щетинки остается полый канал (рис. 4.4.1A).
Поскольку в окончательно сформированной щетинке не остается отростка трихогенной клетки, при очередной линьке [см. Gnathopoda (3b)] щетинка сбрасывается целиком вместе со старой кутикулой, и на сброшенном экзувии все щетинки сохраняют ту же форму и консистенцию, что и на живом животном. Новая щетинка образуется заново на том же месте благодаря функционированию той же трихогенной клетки, которая снова образует дистальный отросток. Перед сбрасыванием старой кутикулы новая щетинка располагается в полости между новой и старой кутикулой тела, не проникая в канал старой щетинки (рис. 4.4.1C). В отличие от кутикулы щетинки, кутикулярный футляр реснички, формируемый текогенной клетки, во время предлиночного процесса не заменяется, а сохраняется и растет в длину. В результате этого сенсорная ресничка вместе со своим удлинившимся футляром проходит сквозь кутикулу новой щетинки и тянется к старой щетинке, так что ресничка имеет два рецепторных участка – один сохраняется на вершине реснички и контактирует со старой щетинкой, а второй образуется в средней части реснички внутри новой щетинки (рис. 4.4.1C).
Отверстие, через которое ресничка выходит из новой щетинки (линочная пора), бывает расположено у основания новой щетинки или на ее вершине (Harris 1977, Zacharuk 1985). При сбрасывании старой кутикулы дистальный участок реснички отрывается, а линочная пора может сохраняться или исчезает. Линочная пора всегда отсутствует на щетинке, появившейся в результате линьки на том месте, где в предыдущем возрасте щетинки не было.
В последнем возрасте жизненного цикла (в частности у имаго Pterygota) в некоторых щетинках трихогенная клетка может отмирать после завершения последнего формирования кутикулы. Некоторые щетинки утратили сенсорную функцию; в этом случае нейрон может не развиваться, а из исходного общего предшественника нейрона и оболочечных клеток развиваются только трихогенная и тормогенная клетки; показано, что так развиваются чешуйки на крыле бабочки (Stossberg 1938) [см. ниже, Lepidoptera (1)] и часть щетинок на крыле ручейника (Rönsch 1954).
(5 c) Разные формы артроподных щетинок. В строении многих щетинок проявляется эволюционная корреляция: щетинки, расположенные в разных местах тела, могут иметь одинаковое или сходное строение, присущее определенному таксону – т.е. в ходе эволюции такие щетинки эволюционировали одновременно. У каждого насекомого может быть много таких типов щетинок, с отдельной эволюционной корреляцией в каждом типе. Некоторые типы щетинок повторяются в родственных таксонах, некоторые являются уникальными. Суммарное число различимых типов щетинок очень велико, и для них нет универсальной номенклатуры. Для описания щетинок используют условную классификацию, в которой в одну категорию относят щетинки сходного строения, которое в некоторых случаях досталось им от общего предка, а во многих случаях возникало независимо в разных филогенетических ветвях.
Наиболее обычной, и, вероятно, исходной формой артроподных щетинок является трихоидная сенсилла (sensillum trichodeum) – это щетинка, постепенно сужающаяся и заостренная на вершине (т.е. оправдывающая название «щетинка»). Основание такой щетинки окружено кольцевидной мембраной [образуемой тормогенной клеткой – см. (5a)], благодаря чему при механическом воздействии щетинка способна отклоняться и выполнять механорецепторную функцию. Кольцевидная мембрана, окружающая основание щетинки, бывает обрамлена жестким кольцевидным кутикулярным возвышением в форме кратера.
Тонкую трихоидную щетинку часто называют волосок а толстую – шиповидной (sensillum chaeticum). Шиповидную щетинку не следует называть «шипом», поскольку этим термином обозначают гиподермосодержащий вырост, не связанный с сенсорными и оболочечными клетками [см. (5d)]. Очень короткую трихоидную сенсиллу называют базиконической (sensillum basiconicum). Многие щетинки не утончаются к концу, а имеют самые разнообразные формы – лопатковидные, булавовидные, зазубренные, гребенчатые, ветвистые, сетчатые и др.
Трихоботрия (trichobothrium) представляет собой тонкую механорецепторную торчащую щетинку, основание которой находится в центре особого кутикулярного образования (например, в центре круглого чашевидного углубления или камеры).
Чешуйка (sensillum squamiformium) представляет собой сильно расширенную уплощенную щетинку с тонким основанием, которое изогнуто так, что чешуйка лежит на покрове. Чешуйки могут покрывать тело, черепицеобразно налегая друг на друга. Чешуйки разнообразного строения имеются у паукообразных (Townsend & Felgenhauer 1998) и насекомых. Среди Hexapoda сходные по строению чешуйки, покрытые многочисленными продольными ребрышками, имеются в таких далеких друг от друга таксонах, как Entognatha, Triplura, Rhipineoptera, Parametabola и Metabola.
Среди Entognatha такие продольно-исчерченные чешуйки имеются у некоторых Diplura-Rhabdura (Lepidocampa/fg и Hemicampa/fg) и у некоторых Collembola-Eupodura (Tomocerus/fg, Lepidocyrtus/fg и др.). Среди Triplura такие же продольно-исчерченные чешуйки покрывают тело у большинства Zygentoma и у всех Microcoryphia [см. ниже, Triplura (4)]. Среди крылатых насекомых покров из таких же чешуек имеется на теле у чешуйчатых сверчков Mogoplistes/fg (см. ниже, Dolichocera: «Классификации» I), на теле и крыльях у некоторых примитивных сеноедов (см. ниже, Parapsocida: «Классификации и статус») и у всех чешуекрылых [см. ниже, Lepidoptera (1)]; такие же по строению чешуйки встречаются у некоторых Trichoptera и Diptera [см. ниже, Pterygota (4Ae)].
Иногда кутикулярная часть щетинки погружена в углубление и/или укорочена так, что не выступает над поверхностью покрова – такое строение имеют целоконическая (sensillum coeloconicum), ампулларная (s. ampullaceum), кампаниформная (s. campaniformium), и плакоидная (s. placodeum) сенсиллы. Иногда кутикулярная часть не выражена – это щелевидные сенсиллы, известные у многих паукообразных и разнообразные сколопофорные сенсиллы, известные у многих насекомых.
Сколопофорная сенсилла (sensillum scolopophorum), или сколопидий (scolopidium) снаружи не отличается от остальной поверхности кутикулы, но внутри тела имеет сенсорные и оболочечные клетки, кутикулярный футляр ресничек и сколопаль. Сколопофорные сенсиллы, собранные пучком и прикрепленные к определенному участку кутикулы в той или иной части тела, образуют хордотональный орган (organum chordotonalium) (рис. 6.6.1B).
На теле животного щетинки могут располагаться строго упорядоченно (так что число и расположение щетинок определенного строения постоянно для вида или надвидового таксона), либо беспорядочно, в этом случае щетинки одинакового строения образуют поля или ряды, в которых число щетинок варьирует индивидуально.
(5 d) Отличие артроподной щетинки от других образований у Euarthropoda. Артроподные щетинки, имеющиеся на покровах Euarthropoda, не следует путать с внешне сходными образованиями иной природы, которые бывают развиты на покровах этих же животных.
Микротрихии и аканты. Микротрихия (microtrichion) или аканта (acantha) – неподвижный кутикулярный вырост, в образовании которого принимает участие лишь одна гиподермальная клетка, тогда как сенсорные и тормогенные клетки отсутствуют. Микротрихией принято называть вырост значительно меньшей толщины, чем гиподермальная клетка; иногда каждая гиподермальная клетка образует лишь одну микротрихию, иногда одна гиподермальная клетка образует много микротрихий. Микротрихию называют также aculeus или aculea. Термин аканта был предложен для более крупного выроста, сопоставимого по толщине с образующей его гиподермальной клеткой (Richards & Richards 1979).
Шпоры и шипы. Подвижная шпора и неподвижный шип – придатки, имеющие внутри многоклеточную гиподерму. По своей форме подвижные шпоры бывают похожи на крупные щетинки; в строении шпор, расположенных в разных местах, может проявляться эволюционная корреляция (подобно эволюционной корреляции у щетинок). При линьке гиподерма, лежащая внутри шпоры, формирует новую кутикулу, так что перед экдизисом новая шпора помещается внутри старой (в отличие от щетинки, которая закладывается вне старой щетинки) (рис. 8.4.4C).
Несмотря на то, что артроподная щетинка и гиподермосодержащая шпора или шип имеют принципиально разное строение, известны случаи, когда они замещают друг друга в онтогенезе или в филогенезе [в частности, см. Pandictyoptera (5) и рис. 8.4.4A–C]. В этих случаях промежуточным состоянием может быть гиподермосодержащий шип, несущий на вершине артроподную щетинку.
Сложные сенсиллы. Сложная щетинкоподобная сенсилла внешне похожа на артроподную щетинку, но содержит не один, а несколько комплектов сенсорных и оболочечных клеток. Каждый такой комплект (или сенсорная единица) может включать несколько сенсорных клеток, две собственные оболочечные клетки (внутреннюю и наружную) и собственную кутикулярную трубку, в которую заключены реснички сенсорных клеток. Кроме того, имеются общие оболочечные клетки, которые окружают все эти наборы клеток. Все кутикулярные трубки с ресничками внутри и окружающие их участки оболочечных клеток заключены в общую кутикулярную оболочку, которая выглядит как одна щетинка. Как и у типичной артроподной щетинки, перикарионы всех клеток, и сенсорных, и оболочечных, расположены не в самой щетинке, а проксимальнее ее, в общем слое гиподермы. Такие сложные щетинкоподобные сенсиллы описаны для Collifera [см. ниже, Diplopoda (1) и Pauropoda (2)] (Nguyen Duy-Jacquemin 1985, Tichy 1987).
(5 e) Отличие артроподной щетинки от щетинок других животных. У некоторых животных, не относящихся к Euarthropoda, имеются образования, также называемые щетинками; эти образования имеют те или иные черты сходства с артроподной щетинкой, но принципиально отличаются от нее.
У других Gnathopoda те же функции механорецепции и хеморецепции выполняют сенсорные выросты, расположенные на разных участках покровов – это сенсорные конусы или циррусы [ср.: Onychophora (3) и Tardigrada (5)]. Как и в артроподных щетинках, внутри этих сенсорных выростов имеются лишь дистальные отростки сенсорных и других клеток, тогда как перикарионы всех клеток лежат в общем слое гиподермы или еще глубже. В отличие от артроподной щетинки, эти сенсорные выросты покрыты тонкой эластичной кутикулой, не образующей склеротизованной щетинки, и при линьке новая кутикула развивается непосредственно под старой кутикулой внутри конуса или цирруса; на сброшенном экзувии на месте сенсорного конуса остается тонкая кутикула, которая может сминаться и не сохранять форму конуса. По крайней мере у Onychophora кутикула конуса вырабатывается не оболочечными клетками, а выростами обычных гиподермальных клеток (Storch & Ruhberg 1993) (рис. 4.2C).
У Nematoidea, как и у Euarthropoda, на толстой внеклеточной кутикуле, сбрасываемой при каждой линьке, имеются щетинки, каждая из которых образована выделениями одной гиподермальной клетки (называемой муфтовой клеткой); муфтовая клетка кольцом окружает дендрит сенсорной клетки. Однако щетинка нематоды имеет следующие отличия от артроподной щетинки: тормогенная клетка отсутствует; муфтовая клетка (которую можно уподобить трихогенной клетке в артроподной щетинке) окружает лишь дистальный участок дендрита; оболочечная клетка, образующая рецепторную полость, является железистой, лежит глубже муфтовой клетки и не участвует в образовании кутикулы; перикарион рецепторной клетки находится не в общем слое гиподермы, а глубже. Щетинки нематод исходно является рецепторно-железистыми образованиями и гомологичны мягким прикрепительным трубочкам, имеющимся на поверхности тела у Gastrotricha (Малахов 1986).
У кольчатых червей (Annelida), плеченогих (Brachiopoda), щетинкочелюстных (Chaetognatha), погонофор (Pogonophora) и некоторых других животных покровы несут аннелидные щетинки (которые также, как и артроподные щетинки, называют щетинками, сетами или хетами); так же, как и артроподные щетинки, аннелидные щетинки состоят из комплекса хитина и белков. Однако животные, обладающие аннелидными щетинками, не имеют хитинсодержащей кутикулы; в отличие от артроподных щетинок, аннелидные щетинки не связаны с сенсорными клетками; каждая аннелидная щетинка образована выделениями преимущественно одной специализированной эпителиальной клетки, погруженной в щетинковый фолликул; аннелидная щетинка пронизана множеством продольных каналов (тогда как артроподная щетинка имеет лишь один центральный канал); к основанию аннелидной щетинки подходят мышцы, обеспечивающие ее активную подвижность (тогда как к артроподной щетинке мышцы никогда не подходят) (Бужинская и др. 1979).
Признаки Euarthropoda неясного филогенетического значения
(6) Антенны. Имеются антенны (antennae), или антенны первые (антенны I); это пара преоральных придатков [см. Metameria (3)], расположенных на голове и иннервируемых от надглоточного ганглия; в связи с этим надглоточный ганглий разделен на два отдела: протоцеребрум (protocerebrum) – передний отдел, иннервирующий зрительные органы [глазки и глаза – см. (4)], и дейтоцеребрум (deutocerebrum) – задний отдел, иннервирующий антенны. Дейтоцеребрум соединен непосредственно с посторальным отделом нервной системы; при этом первый посторальный ганглий сдвинут дорсально и плотно соединен с надглоточным ганглием, в связи с чем считается частью надглоточного ганглия и называется тритоцеребрум (tritocerebrum). Обычно антенны расположены впереди глаз; антеннальные нервы отходят от вентральной (или передней) стороны дейтоцеребрума и идут вперед к антеннам, обходя протоцеребрум и его зрительные доли с вентральной стороны. Так что судя по иннервации, антенны являются исходно вентральными придатками, но у различных членистоногих занимают вентральное, дорсальное, латеральное или переднее положение. Антенна обычно состоит из некоторого числа члеников [см. (1)] – антенномеров (antennomeri, или antennomera); к основанию первого антенномера подходят мышцы, крепящиеся в голове, а в каждом антенномере имеются мышцы, идущие к основанию следующего членика. В разных таксонах число антенномеров различается, бывает определенным или неопределенным, меняется или не меняется в онтогенезе (см. Указатель признаков [1.28] и табл. B).
Некоторые авторы считают антенны I гомологами посторальных конечностей; однако у дейтоцеребрума, как и у протоцеребрума, левая и правая половины соединены между собой комиссурой, проходящей впереди (дорсальнее) пищевода, так что иннервируемые им антенны I относятся к преоральной области.
Возможно, нечленистые предшественники антенн исходно имеются у Metameria. У онихофор имеется пара преоральных придатков, считающихся гомологичных антеннам [ср. Onychophora (4)]. Две или более пар преоральных придатков имеются у Polychaeta; возможно, одна из этих пар – пальпы или щупальца – гомологичны антеннам членистоногих. Некоторые авторы ошибочно принимали за гомологи антенн циррусы тихоходок [см. (5d); ср.: Tardigrada (5)].
Среди современных членистоногих антенны имеются только у Mandibulata, тогда как у хелицеровых антенны всегда отсутствуют [см. ниже, Chelicerophora (1)]. Помимо Mandibulata, антенны имеются у Trilobitomorpha; поскольку все Trilobitomorpha известны лишь по ископаемым остаткам, иннервация их антенн неизвестна. Так что не вполне ясно, является ли наличие настоящих преоральных антенн аутапоморфией Euarthropoda, или это лишь аутапоморфия Mandibulata.
Антенны ракообразных принято называть антенны I (антенны первые) или антеннулы (antennulae)], поскольку помимо них имеются еще антенны II (антенны вторые), которые иннервируются от тритоцеребрума и являются конечностями I посторального сегмента [см. ниже, Eucrustacea (2) и (6)]. У насекомых и многоножек антенны преоральной области являются единственными [см. ниже, Atelocerata (1)], поэтому их называют просто антенны, или усики (в старой литературе – сяжки).
Среди Mandibulata антенны вторично утрачены у некоторых Eucrustacea, у Protura и у некоторых крайне специализированных представителей Pterygota (это личинки и самки некоторых Gallinsecta и Strepsiptera, ведущие неподвижный образ жизни).
Следует упомянуть, что у некоторых членистоногих имеются парные придатки неясного происхождения, лежащие впереди антенн I и называемые преантенны. У взрослых животных преантенны имеются у некоторых ракообразных – у всех Remipedia [см. ниже, Remipedia (2)] и у нескольких средиземноморских видов Eucrustacea-Malacostracia-Isopoda-Jaera/g (Verhoeff 1943); в эмбриогенезе преантенны обнаружены у некоторых Chilopoda и Hexapoda (Snodgrass 1938, Иванов 1940).
(7) Верхняя губа. Имеется верхняя губа (labrum) – непарный выступ или складка, свисающая впереди ротового отверстия. Некоторые авторы считают, что образования, называемые «верхней губой» у различных членистоногих – у Chelicerata, Eucrustacea и Atelocerata, не гомологичны между собой. Согласно противоположной крайней точке зрения верхняя губа исходно имеется у Gnathopoda; в этом случае за гомолог верхней губы у онихофор принимают дорсальную складку на валике, образующемся вокруг челюстей при их втягивании в голову [ср.: Onychophora (1)] (Butt 1959).
Геологический возраст. Кембрий – ныне (рис. 2.2).
Систематическое положение Euarthropoda. Помимо принятого здесь отнесения Euarthropoda непосредственно к Gnathopoda, некоторые авторы объединяют Euarthropoda и Tardigrada в таксон Arthropoda (см. Gnathopoda: «Классификации» II).
Статус Euarthropoda. Некоторые авторы не признают таксон Euarthropoda (см. Gnathopoda: «Классификации» I).
Классификации Euarthropoda. Из многих предлагавшихся классификаций членистоногих рассмотрим наиболее известные.
Таксон Branchiata Lang 1888 {syn.circ.: Carides Haeckel 1866; по циркумскрипции соответствует: Crustacea auct. (non Brünnich 1771, nec Pennant 1777} включает всех первичноводных членистоногих (Eucrustacea, Trilobita, Xiphosura и др.), и характеризуется только плезиоморфиями – наличием жаберного дыхания и сохранением выделительной системы в виде целомодуктов [см. Peritoneata (1)]. Это заведомо парафилетический таксон, предковый для Tracheata.
Таксон Tracheata Haeckel 1866 {syn.circ.: Entomia Rafinesque 1815; по циркумскрипции соответствует: «Insecta» sensu Latreille 1806 (non Linnaeus 1758)} объединяет Atelocerata и Arachnida; исходно Геккель причислял сюда также Pantopoda и Tardigrada, в то время считавшихся паукообразными, а позже (Haeckel 1889) причислил сюда Onychophora. Таксон Tracheata считался голофилетическим, произошедшим одной ветвью от Branchiata, в пользу чего свидетельствуют две апоморфии, связанные с переходом этих животных к наземному образу жизни: (1) наличие трахейной системы и (2) наличие мальпигиевых сосудов. Однако после того, как были описаны синапоморфии Xiphosura и Arachnida, стало ясно, что Tracheata – это полифилетический таксон, и его признаки не могут быть однократно возникшими апоморфиями (см. II и рис. 4.4.2B). В результате критического исследования первого из этих признаков, было выявлено, что трахейная система у разных Tracheata образуется из впячиваний покровов в принципиально разных местах тела, из чего следует, что сходная по строению трахейная система независимо возникла у Hexapoda, Symphyla, Collifera, Notostigmophora, Pleurostigmophora и различных Arachnida [см. Gnathopoda (3c)]. В результате критического исследования второго признака, считавшегося апоморфией Tracheata, выяснилось, что, несмотря на то, что мальпигиевы сосуды у Atelocerata и Arachnida имеют одинаковое строение и функцию, они закладываются из разных участков кишечника и образованы разными зародышевыми листками: у Arachnida мальпигиевы сосуды отходят от средней кишки и, таким образом, являются энтодермальными, а у Atelocerata они отходят от границы средней и задней кишки и построены из материала задней кишки, т.е. являются эктодермальными [см. ниже, Atelocerata (7)]. Таким образом, из теории зародышевых листков, утверждающей, что апоморфией Monostomata является необратимая дифференциация эктодермы и энтодермы, следует, что мальпигиевы сосуды Atelocerata и Chelicerata не являются гомологичными органами.
Следующие две классификации основаны на гипотезе о монофилии Euarthropoda (о противоположной точке зрения – см. Gnathopoda: «Классификации» I). Согласно этой гипотезе общим предком всех Euarthropoda были морские членистоногие с большим неопределенным числом сегментов тела и гомономными полифункциональными конечностями, подобно тому, что имелось у вымерших палеозойских Trilobitomorpha. Каждая конечность выполняла и жевательную функцию (благодаря наличию на коксоподите жевательного выступа – гнатобазы), и локомоторную функцию (за счет развитого членистого дистального отдела – телоподита), и дыхательную функцию (за счет наличия жаберного отростка) [см. Euarthropoda (3)]. Дальнейшая эволюция членистоногих шла по пути дифференциации конечностей и соответствующих сегментов тела, в связи с чем образуются отделы тела – тагмы [см. Metameria (1c)]. Поскольку дифференциация конечностей и соответствующих тагм происходила независимо в разных филогенетических ветвях Euarthropoda, сегментарный состав их тагм разный (табл. IV.1 и рис. 4.1).
II. Деление Euarthropoda на Teleiocerata, Chelicerophora и Atelocerata (рис. 4.4.2C). Таксон Teleiocerata Heymons 1901 включает Eucrustacea и Trilobita и является плезиоморфоном, предковым и для голофилетического таксона Chelicerata Heymons 1901, и для голофилетического таксона Atelocerata Heymons 1901. Исходно эти таксоны были охарактеризованы по специализации сегментов, вошедших в состав головы. Антенны первые (т.е. преоральные придатки по принятой здесь терминологии, или придатки II метамера по терминологии Хаймонса) сохраняются у Teleiocerata и Atelocerata, но утрачены у Chelicerata. Антенны вторые (т.е. придатки I посторального сегмента по принятой здесь терминологии, или III метамера по терминологии Хаймонса) сохраняются у Teleiocerata и преобразованы в клешни (хелицеры) у Chelicerata, но утрачены у Atelocerata (рис. 4.1). Таким образом, таксон Teleiocerata характеризуется только плезиоморфией (сохранением обеих пар антенн), a таксоны Chelicerata и Atelocerata характеризуются каждый своими аутапоморфиями (Chelicerata – утратой антенн I и преобразованием антенн II, Atelocerata – утратой антенн II). В настоящее время к этим апоморфиям добавлены другие, и голофилетический таксон Chelicerata признан в составе таксона Chelicerophora, а голофилетический таксон Atelocerata признан в составе таксона Mandibulata [см. ниже, Chelicerophora (1)–(3) и Atelocerata (1)–(3)].
![]() Таблица
IV.1. Специализация головных и
головогрудных сегментов у Atelocerata, Chelicerata и
Telieocerata
Таблица
IV.1. Специализация головных и
головогрудных сегментов у Atelocerata, Chelicerata и
Telieocerata
|
По Heymons 1901: |
||||||
|
|
Atelocerata |
Chelicerata |
Teleiocerata |
|||
|
Myriapoda (Scolopendra) |
Insecta (Forficula) |
Arachnoiden (Scorpio) |
Gigantostraca (Limulus) |
Trilobita (Triarthrus) |
Crustacea (Branchipus) |
|
|
акрон |
||||||
|
1.сегмент |
||||||
|
2.сегмент |
антенны |
антенны |
хелицеры |
хелицеры |
антенны |
1.антенны |
|
3.сегмент |
– |
– |
педипальпы |
1.гнатоподы |
1.гнатоподы |
2.антенны |
|
4.сегмент |
мандибулы |
мандибулы |
1.ноги |
2.гнатоподы |
2.гнатоподы |
мандибулы |
|
5.сегмент |
1.максиллы |
1.максиллы |
2.ноги |
3.гнатоподы |
3.гнатоподы |
1.максиллы |
|
6.сегмент |
2.максиллы |
2.максиллы |
3.ноги |
4.гнатоподы |
4.гнатоподы |
2.максиллы |
|
7.сегмент |
|
|
4.ноги |
5.гнатоподы |
|
|
|
То же, но с современной гомологизацией и исправленными названиями: |
||||||
|
|
Atelocerata |
Chelicerata |
Teleiocerata |
|||
| Myriapoda |
Hexapoda |
Arachnida |
Xiphosura |
Trilobita |
Eucrustacea |
|
|
преоральн. |
антенны |
антенны |
– |
– |
антенны |
1.антенны |
|
1.сегмент |
– |
– |
хелицеры |
хелицеры |
1.гнатоподы |
2.антенны |
|
2.сегмент |
мандибулы |
мандибулы |
педипальпы |
1.гнатоподы |
2.гнатоподы |
мандибулы |
|
3.сегмент |
(разное) |
максиллы |
1.ноги |
2.гнатоподы |
3.гнатоподы |
1.максиллы |
|
4.сегмент |
(разное) |
лабиум |
2.ноги |
3.гнатоподы |
|
(разное) |
|
5.сегмент |
|
|
3.ноги |
4.гнатоподы |
|
|
|
6.сегмент |
|
|
4.ноги |
5.гнатоподы |
|
|
III. Деление Euarthropoda на Trilobitomorpha, Chelicerophora и Mandibulata (рис. 4.4.2E). Дальнейшее изменение классификации было предложено Снодграссом (Snodgrass 1935, 1938a), расформировавшим парафилетический таксон Teleiocerata за счет создания голофилетического таксона Mandibulata (рис. 4.4.2D). В этой классификации сохранены голофилетические таксоны Atelocerata и Chelicerata, но исчез крупный парафилетический таксон Teleiocerata. Таксон Chelicerata объединен с Pantopoda в таксон Chelicerophora (который в публикациях Снодграсса фигурирует под названием «Chelicerata»). Сравнение сегментарного состава тагм у Chelicerophora и Mandibulata дано в табл. IV.2.
Интересно отметить, что такое же деление членистоногих на Pseudognatha и Polygnatha (приблизительно соответствующих по циркумскрипции Chelicerophora и Mandibulata) за сто с лишним лет до Снодграсса предлагал в одной из своих работ Латрейль (Latreille 1820) (см. ниже, Chelicerophora: «Название» и Mandibulata: «Название»). Однако позже Латрейль вернулся к традиционному делению членистоногих на первичноморских и наземных (см. I).
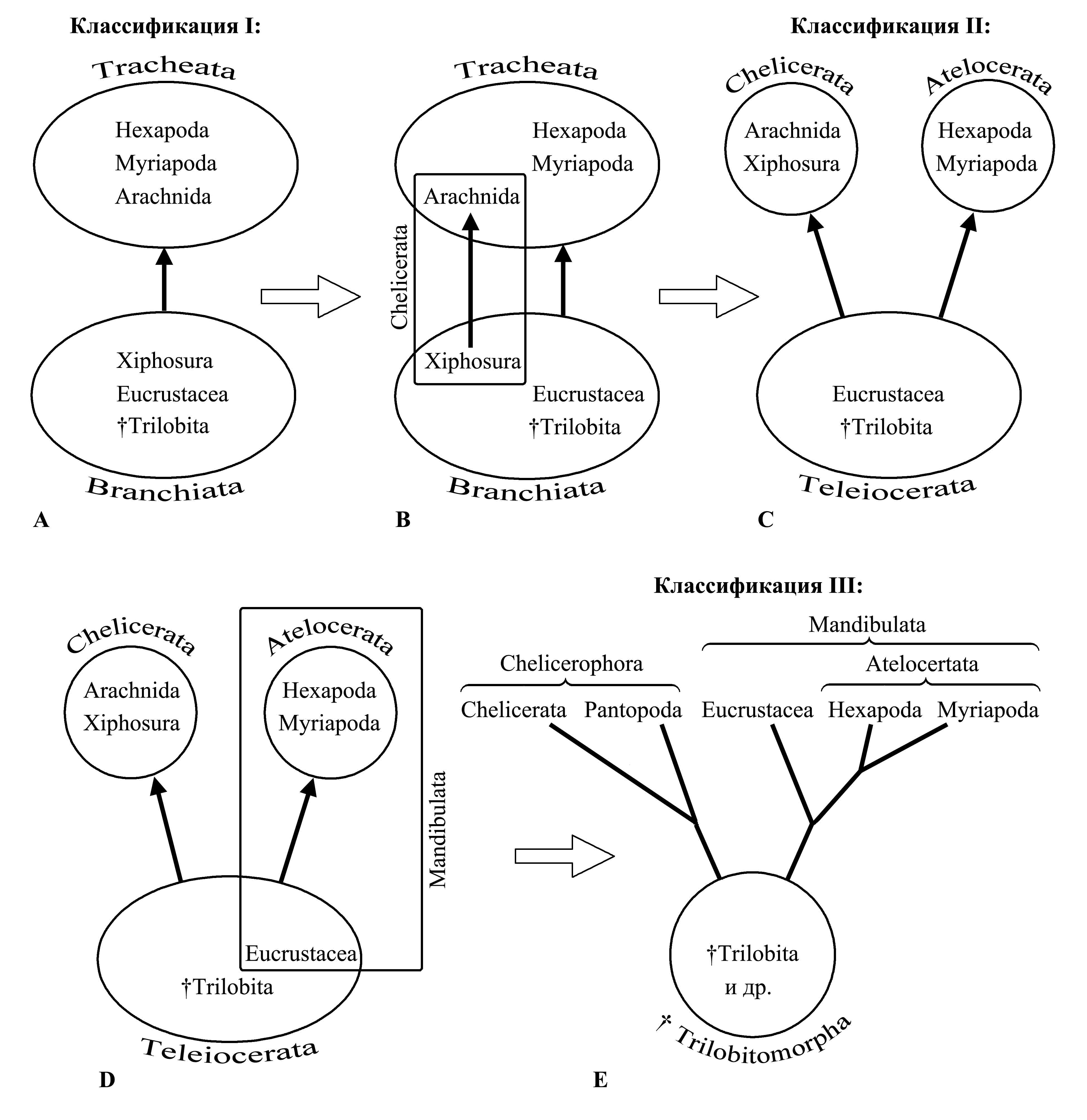
Рис. 4.4.2. Изменения классификации членистоногих в результате уточнения и исправления представлений об их филогении. Черными линиями обозначены филогенетические ветви, стрелками – направления эволюции; светлыми стрелками обозначены изменения классификации. Номера классификаций соответствуют номерам в разделе «Классификации Euarthropoda». A – традиционная классификация (Haeckel 1866 и др.); B – выявление полифилии Tracheata благодаря обнаружению синапоморфий Xiphosura и Arachnida (Lankester 1881); C – классификация с учетом синапоморфий Xiphosura и Arachnida (Heymons 1901); D – выявление синапоморфий Eucrustacea и Atelocerata; E – классификация с учетом синапоморфий Eucrustacea и Atelocerata (Snodgrass 1935).
![]() Таблица
IV.2.
Специализация сегментов тела и их
конечностей у Chelicerophora и Mandibulata
Таблица
IV.2.
Специализация сегментов тела и их
конечностей у Chelicerophora и Mandibulata
|
|
Chelicerophora |
Mandibulata |
||||
|
Pantopoda |
Chelicerata |
|||||
|
Придатки: |
Придатки: |
Тагмы: |
Придатки: |
Тагмы: |
||
|
Преоральный |
– |
– |
головогрудь |
антенны
I |
голова |
|
|
Пост- |
I |
хелифоры |
хелицеры |
антенны
II |
||
|
II |
педипальпы |
гнатоподы,
ноги |
мандибулы |
|||
|
III |
овигеры |
гнатоподы
или ноги |
максиллы
I |
|||
|
IV |
ноги |
гнатоподы
или ноги |
конечности, |
|||
|
V |
ноги |
гнатоподы
или ноги |
туловище |
|||
|
VI |
ноги |
гнатоподы
или ноги |
||||
|
VII |
ноги |
хилярии |
|
|||
|
VIII и |
– |
половые |
брюшко |
|||
Здесь мы принимаем в составе Euarthropoda три подчиненных таксона – Trilobitomorpha, Chelicerophora (стр. 000) и Mandibulata (стр. 000).
Euarthropoda, или Araneus/fg7
![]() 1. Плезиоморфон Trilobitomorpha,
или Calymene/fg1
1. Плезиоморфон Trilobitomorpha,
или Calymene/fg1
![]() 2. Chelicerophora,
или Araneus/fg8
2. Chelicerophora,
или Araneus/fg8
![]() 2.1. Pantopoda,
или Pycnogonum/fg1
2.1. Pantopoda,
или Pycnogonum/fg1
![]() 2.2. Chelicerata,
или Araneus/fg9.
2.2. Chelicerata,
или Araneus/fg9.
![]() 3. Mandibulata,
или Scarabaeus/fg1
3. Mandibulata,
или Scarabaeus/fg1
![]() 3.1. Eucrustacea, или Cancer/fg1
3.1. Eucrustacea, или Cancer/fg1
![]() 3.2. Atelocerata, или
Scarabaeus/fg2
3.2. Atelocerata, или
Scarabaeus/fg2
![]() 3.2.1. Myriapoda, или
Scolopendra/fg1
3.2.1. Myriapoda, или
Scolopendra/fg1
![]() 3.2.
2. Hexapoda, или Scarabaeus/fg3
3.2.
2. Hexapoda, или Scarabaeus/fg3