КЛАДОЭНДЕЗИС НАСЕКОМЫХ

![]()
|
КЛАДОЭНДЕЗИС НАСЕКОМЫХ |
|||||||
|
|
|
fg | |||||
IV-1.3.3.2. Atelocerata Heymons 1901 – неполноусые
|
Названия, однозначно соответствующее по циркумскрипции: — Dicera Latreille 1802a; — Antennata sensu Lang 1888 (non sensu Börner 1909; nec Antennata Leunis 1860); — Atelocerata Heymons 1901; — Eutracheata Grobben 1908; — Monocerata Ulrich 1951. По циркумскрипции также соответствует: — «Insectes» sensu Cuvier 1817 (non Insecta Linnaeus 1758); — «Tracheata» sensu Pocock 1893a (non Haeckel 1866). Иерархическое название: Pediculus/f2=Scarabaeus/g2 [f:1777; g:L.1758(gen.170)] (sine Cancer; incl. Scolopendra). |
|
Ранг. Обычно от надкласса до подтипа.
Название. Часто применяемое для этого таксона название «Tracheata» не соответствует ему по исходной циркумскрипции. Исходно (Haeckel 1866) название Tracheata применялось к подтипу членистоногих, включавшему, помимо Hexapoda и Myriapoda, также Arachnida; этот подтип, считавшийся монофилетическим, на самом деле представляет собой полифилетический таксон, происходящий не менее чем двумя ветвями от подтипа Carides, или Branchiata (см. Euarthropoda: «Классификации» I).
Название Antennata, возможно, является старейшим названием, соответствующим по циркумскрипции обсуждаемому ниже таксону; однако это же название используется в литературе и в других значениях, а истинное авторство названия Antennata не выяснено. Здесь мы используем для таксона, объединяющего Myriapoda и Hexapoda, название Atelocerata, как бесспорно соответствующее по циркумскрипции и всегда употреблявшееся только в этом значении.
Аутапоморфии Atelocerata
(1) Интеркалярный сегмент. Утрачены конечности первого посторального сегмента (соответствующие антеннам вторым Eucrustacea и хелицерам Chelicerophora – см. Указатель признаков [0.7]). Таким образом, имеется лишь одна пара антенн, являющихся преоральными придатками [см. Euarthropoda (6)] и соответствующих антеннам первым Eucrustacea (с чем связано название «Atelocerata», или «неполноусые»); эта единственная пара антенн развита у большинства Atelocerata и утрачена только у Protura и некоторых крайне специализированных Pterygota.
Поскольку у Atelocerata первый посторальный сегмент не несет конечностей и внешне не выражен, его принято называть интеркалярным (т.е. вставочным). Об исходном существовании этого сегмента у предков Atelocerata свидетельствует наличие принадлежащих ему сегментарных органов – это тритоцеребрум [ганглий I посторального сегмента – см. Euarthropoda (6)], имеющийся у всех неполноусых, и целомические мешки и/или зачатки конечностей I посторального сегмента, закладывающиеся в эмбриогенезе у некоторых неполноусых [см. Gnathopoda (1)]. Тритоцеребрум обычно выглядит как пара утолщений в местах отхождения окологлоточных коннектив от надглоточного синганглия (рис. 6.6.2C). Тритоцеребрум является ясно обособленным ганглием: он удален от подглоточного синганглия (который является результатом слияния ганглиев II–IV посторальных сегментов) и слит с надглоточным синганглием (головным мозгом); в то же время от остальных отделов надглоточного синганглия (протоцеребрума, иннервирующего глаза, и дейтоцеребрума, иннервирующего антенны) тритоцеребрум отличается тем, что его комиссура проходит под пищеводом, т.е. он является самым передним ганглием посторального происхождения. Тритоцеребрум иннервирует верхнюю губу и углы рта – т.е. область впереди мандибул (придатков II посторального сегмента), а также органы Темешвари [см. (2)].
(2) Органы Темешвари. На голове исходно имеется пара височных, или темпоральных органов (organa temporalia), называемых также постантеннальные органы, или органы Темешвари (Tömösváry 1883). Это хеморецепторные органы, образованные пористой кутикулой и расположенными под ней ольфакторными клетками. Орган Темешвари расположен позади основания антенны, но впереди сложного глаза [см. Euarthropoda (4)]. Органы Темешвари иннервируются от тритоцеребрума и, возможно, являются производными исчезнувших антенн вторых [см. (1)].
Органы Темешвари имеются у многих Myriapoda и многих Entognatha, но отсутствует у всех Amyocerata. Среди Myriapoda они развиты у Symphyla (рис. 5.2.1D), Pauropoda (рис. 5.4.1B), примитивных Diplopoda (Penicillata, Opisthandria, Nematophora, Merocheta) и примитивных Chilopoda (Notostigmophora, Ungulipalpi, Craterostigmus/fg) (рис. 5.5.2D), но утрачены у части Diplopoda и части Chilopoda (Devonobius/fg и все Epimorpha). Среди Entognatha органы Темешвари имеются у некоторых Diplura, многих Collembola (рис. 6.4.1A) и всех Protura [ложные глазки – см. ниже, Protura (1) и рис. 6.5B].
Строение органов Темешвари у разных Atelocerata разнообразно. Некоторые авторы считают их гомологичными у всех Atelocerata (Haupt 1979); по мнению других авторов они могли независимо возникать у разных Atelocerata, и в таком случае их наличие не является синапоморфией.
(3) Претарсус. Самый дистальный членик ноги [см. Euarthropoda (3)] – претарсус (praetarsus), называемый также коготком (unguis), или дактилоподитом (от «dactylo-» и «pod-»). Он не имеет мышц-разгибателей; от основания претарсуса вглубь ноги отходит единственная аподема – унгуитрактор (unguitractor), к которой крепятся все мышцы, сгибающие претарсус (унгуитрактором также называют обособленный склерит в основании претарсуса, от которого отходит эта аподема); аподема унгуитрактора и отходящие от нее мышцы проходят сквозь предыдущий отдел ноги – лапку, или тарсус (tarsus), так что неподвижные концы этих мышц крепятся не в тарсусе, а в более проксимальных члениках ноги; таким образом, внутри тарсуса нет мест прикрепления мышц (рис. 6.1.2). Тарсус бывает одночлениковым или разделен несколько безмышечных члеников. В отличие от Atelocerata, у других членистоногих коготки ног могут иметь не только мышцы-сгибатели, но и мышцы-разгибатели, а неподвижные концы этих мышц могут крепиться в разных члениках ноги, включая членик, к которому крепится коготок.
Признаки Atelocerata неясного филогенетического значения
(4) Утрата телоподита мандибулы. Мандибула [см. Mandibulata (1)] без щупика, т.е. состоит из единственного членика – коксоподита. Неуникальная апоморфия, так как редукция мандибулярного щупика происходит также у различных Eucrustacea. Некоторые авторы расценивали отсутствие мандибулярного щупика как первичное и считали это доводом в пользу теории о полифилии членистоногих [см. Mandibulata (1)]. У многоножек единственный членик мандибулы, ее коксоподит, имеет усложненное строение, с подвижно соединенными склеритами, что послужило поводим считать их мандибулы состоящими более чем из одного членика [см. ниже, Myriapoda (1)].
(5) Конечности одноветвистые. Конечности неразветвленные [в отличие от двуветвистых у ракообразных – ср.: Eucrustacea (2) и рис. 4.7.1F]. Существует предположение, что стилусы около оснований ног у Symphyla (рис. 5.2.1D–E) и стерностили на брюшке у насекомых [см. ниже, Hexapoda (7) и рис. 6.8.1C] являются вестигиями второй ветви конечности, имевшейся у общих предков Atelocerata и Eucrustacea. Возможно, двуветвистое строение сохраняется у максилл насекомых [см. ниже, Hexapoda (2)]. В противоположность этому приверженцы полифилетической теории происхождения членистоногих (см. Gnathopoda: «Классификации» I) считали, что конечности Atelocerata произошли из ног Onychophora и поэтому исходно одноветвисты (в связи с чем для таксона Ceratophora, включающего Atelocerata и Onychophora, было предложено название Uniramia).
(6) Среда обитания. Большинство неполноусых ведут наземный образ жизни; лишь некоторые насекомые вторично перешли к водному, преимущественно пресноводному, образу жизни [см. ниже, Hexapoda (9)]. С наземным образом жизни связано наличие мальпигиевых сосудов [см. (7)] и трахейной системы. Трахейная система [независимо возникающая у разных членистоногих – см. Gnathopoda (3c)] имеется у большинства неполноусых. Вероятно, она возникла в разных таксонах неполноусых независимо, о чем свидетельствует разное положение трахеостий: у Chilopoda-Pleurostigmophora и у большинства Hexapoda имеется по паре трахеостий (обычно в виде дыхалец) на боках туловищных сегментов; у Collifera трахеи открываются в коксальные аподемы, а у Symphyla – в мандибулярные аподемы; у Collembola дыхальца находятся на шее; у Chilopoda-Notostigmophora непарные дыхальца находятся на тергитах туловища. У некоторых неполноусых – у Pauropoda-Tetramerocerata, у многих Collembola и многих Protura – трахейная система отсутствует. Высказано предположение, что исходно неполноусые имели трахейную систему с дыхальцами на плейритах, которые сохранились у Pleurostigmophora и Hexapoda и сместились на коксальные аподемы у Collifera (Klass & Kristensen 2001).
Известны ископаемые остатки, относимые к многоножкам и датируемые силурийским периодом (см. ниже, Diplopoda: «Геологический возраст» и Notostigmophora: «Геологический возраст»). На основании представления о том, что в силуре не было наземных биоценозов, высказано предположение, что эти многоножки были первичноморскими; в этом случае наземный образ жизни современных многоножек и насекомых и наличие у них трахейной системы является не аутапоморфией Atelocerata, а результатом конвергенции. Однако имеются данные о существовании в силуре наземных биоценозов (см. главу II.2); это позволяет предположить, что силурийские многоножки, как и современные, обитали на суше.
(7) Мальпигиевы сосуды. Имеются мальпигиевы сосуды (исходно обозначенные как сосуды «N» и «P» в книге Malpighi 1687, Tom II, Tab. V, Fig. I). Мальпигиевы сосуды представляют собой трубчатые выросты эктодермального происхождения, отходящие от кишечника на границе средней (энтодермальной) и задней (эктодермальной) кишки [см. Haemataria (1)] и слепо заканчивающиеся в полости тела. Мальпигиевы сосуды – выделительные органы; у Atelocerata они функционально заменили целомодукты и их производные, служащие выделительными органами у других членистоногих и у кольчатых червей [см. Peritoneata (1) и Gnathopoda (1)]. В отличие от целомодуктов, мальпигиевы сосуды выводят экскреты не наружу, а в кишку; при прохождении экскретов по кишечнику содержащаяся в них вода всасывается ректальными железами задней кишки. В результате, наружу выделяются не жидкие, а твердые экскреты – преимущественно мочевая кислота, что является одной из важных адаптацией Atelocerata к жизни на суше [см. (6)]; у вторичноводных Atelocerata выделяются жидкие экскреты – аммиак и др. У личинок некоторых насекомых (Ephemeroptera-Asthenopus/fg, некоторые Thysanoptera, некоторые Eleuterata, Birostrata, Rhaphidioptera) мальпигиевы сосуды выделяют шелк, выводящийся из анального отверстия и используемый для сооружения убежища (у других насекомых шелк выделяется железами иного происхождения – см. Указатель признаков [4.3.2]). Мальпигиевы сосуды имеются во всех группах Atelocerata, но у многих Entognatha они имеют вид маленьких нефункционирующих вестигиев. Число мальпигиевых сосудов бывает различным – от двух (одной пары) до неопределенно большого числа; в строении мальпигиевых сосудов разных пар проявляется эволюционная корреляция. Иногда мальпигиевы сосуды утрачены (см. Указатель признаков [4.7]: табл. F). У некоторых насекомых имеется криптонефрия: дистальный конец мальпигиевого сосуда не оканчивается свободно, а соединяется с эпителием задней кишки, образуя криптонефридий (cryptonephridium). Во всех случаях полость мальпигиева сосуда сообщается с полостью кишки только в месте первичного отхождения сосуда на границе средней и задней кишки; в месте вторичного соединения мальпигиевого сосуда с задней кишкой вещества из полости сосуда могут попадать в полость кишки лишь путем диффузии сквозь эпителий сосуда и эпителий кишки.
У Myriapoda обычно имеется 2 мальпигиевых сосуда (т.е. 1 пара). У Entognatha мальпигиевы сосуды редуцированы: у Diplura они либо совсем отсутствуют, либо имеют вид коротких полых папилл, число которых у разных видов бывает от 5 до 16; у Collembola на месте мальпигиевых сосудов может быть лишь утолщение кишечного эпителия; у Protura имеется 6 мальпигиевых органов, каждый их которых состоит лишь из двух клеток. У Zygentoma 4–8 сосудов (т.е. 2–4 пары). У Microcoryphia 12–20 сосудов. У личинок Ephemeroptera имеется неопределенно большое число мальпигиевых сосудов особого строения [см. ниже, Ephemeroptera (16)]. У Odonata 20–70 сосудов. У Embioptera 20–30 сосудов. У Notoptera 14–20 сосудов. У Plecoptera 20–100 сосудов. У Raptoriae около 100 сосудов. У Neoblattariae 80–120 сосудов, собранных в 6 групп. Среди термитов у Hemiclidoptera 12–15 сосудов; у Cryptoclidoptera число сосудов уменьшено и стабилизировано, не превышает 8 штук, из которых только 4 закладываются в эмбриогенезе; среди них у Metatermitidae лишь 2–4 сосуда (см. ниже, Isoptera: «Классификация»). У Saltatoria до 300 сосудов. У Spectra несколько десятков сосудов. У Dermatoptera 8–20 сосудов. У Eumetabola число мальпигиевых сосудов уменьшено и стабилизировано, не превышает 8 штук (т.е. 4 пар) [см. ниже, Eumetabola (5)].
Независимо от Atelocerata аналогичные органы выделения (также называемые мальпигиевыми сосудами) возникли у Arachnida – наземной группы в составе Chelicerata, а также у некоторых наземных Eucrustacea; но в отличие от мальпигиевых сосудов Atelocerata выделительные органы этих животных открываются в среднюю кишку и имеют энтодермальное происхождение. Согласно теории зародышевых листков, утверждающей, что апоморфией Monostomata является необратимая дифференциация зародыша на эктодерму и энтомезодерму [см. Monostomata (1)], мальпигиевы сосуды Atelocerata и Chelicerata, образующиеся за счет разных зародышевых листков, не могут быть гомологичными органами и, следовательно, имеют независимое происхождение. Мальпигиева сосуды эктодермального происхождения, помимо Atelocerata, имеются у некоторых тихоходок [ср.: Tardigrada (9)], где они, вероятно, также возникли независимо.
Геологический возраст. Палеозой – ныне. Достоверно известны начиная с девона (см. ниже, Pleurostigmophora и Collembola). По некоторым данным существовали в силуре (см. ниже, Diplopoda и Notostigmophora).
Систематическое положение Atelocerata. Помимо принятого здесь отнесения Atelocerata к Mandibulata, предлагались объединять Atelocerata и Onychophora в таксон Ceratophora (см. Gnathopoda: «Классификации» I).
Статус Atelocerata. Некоторые авторы не признают таксон Atelocerata; одной из версий, альтернативных предположению о голофилии Atelocerata, является предположение о голофилии Pancrustacea (см. Mandibulata: «Классификации» II).
Классификации Atelocerata
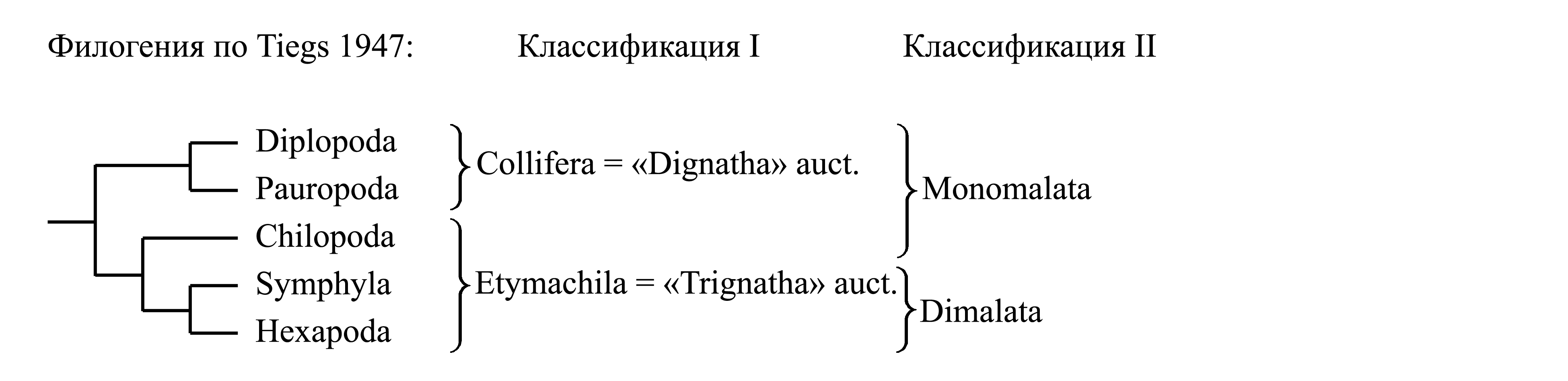
I. Деление Atelocerata на «Dignatha» auct. и «Trignatha» auct. Эта классификация основана на предположении, что в ходе эволюции неполноусых в состав головы последовательно включались лежащие позади нее сегменты туловища, и при этом добавление нового сегмента происходило однократно.
Таксон Collifera, по циркумскрипции соответствующий «Dignatha» sensu Lankester 1904, объединяет Diplopoda и Pauropoda и характеризуется тем, что в ротовом аппарате имеется лишь 2 пары конечностей, так что голова включает лишь 3 посторальных сегмента (рис. 4.1) [см. ниже, Collifera (1) и (2)].
Таксон Etymachila Bollman 1893 [по циркумскрипции соответствует: «Trignatha» sensu Tiegs 1947 (non Lameere 1895)] объединяет Chilopoda, Symphyla и Hexapoda и характеризуется тем, что в ротовом аппарате имеется 3 пары конечностей, так что голова включает 4 посторальных сегмента (рис. 4.1). Этот признак рассматривался как аутапоморфия, на основании чего таксон Etymachila считали голофилетическим. Однако, по-видимому, у Chilopoda превращение конечностей IV посторального сегмента в ротовые придатки (щупики) произошло независимо от других неполноусых [см. ниже, Chilopoda (3)].
Конечности этого же сегмента явно независимо специализировались как ротовые придатки также у большинства Eucrustacea, о чем свидетельствует отсутствие их специализации у цефалокарид [ср.: Cephalocarida (4)].
II. Деление Atelocerata на Monomalata и Dimalata.
Таксон Monomalata Sharov 1966 объединяет Collifera и Chilopoda и характеризуется тем, что лишь одна пара челюстей является жующей — это мандибулы [см. ниже, Myriapoda (1)], тогда как следующая (III посторальная) пара конечностей слилась в непарное лабиумоподобное образование [см. ниже, Collifera (1) и Chilopoda (2)] (рис. 4.1).
Таксон Dimalata Sharov 1966 {по циркумскрипции соответствует: «Labiata» sensu Tiegs 1947 (non Snodgrass 1938a) = «Insecta» sensu Packard 1883 (non Linnaeus 1758)} объединяет Symphyla и Hexapoda и характеризуется тем, что две пары челюстей являются жующими – это мандибулы и максиллы [см. ниже, Symphyla (3) и Hexapoda (2)] (рис. 4.1). Названия Monomalata и Dimalata не следует путать со сходными по этимологии названиями Monognatha, Dignatha и Trignatha: в первом случае речь идет о числе грызущих пар челюстей, а во втором – об общем числе пар придатков в ротовом аппарате.
Выделение таксона Dimalata связано с симфильной теорией происхождения насекомых – т.е. с представлением о родстве Symphyla и Hexapoda (см. ниже, Symphyla: «Систематическое положение»).
. В подтверждение этой теории приводят целый набор общих признаков Symphyla и Hexapoda:
(1) Одинаковый набор ротовых придатков: пара мандибул, пара максилл и непарная нижняя губа [см. ниже, Symphyla (3), (4) и Hexapoda (2), (3)].
(2) Наличие стилусов и расположенных вентральнее их втяжных пузырьков на сегментах туловища [см. ниже, Symphyla (6) и Hexapoda (7)].
(3) Наличие на заднем крае последнего тергита туловища пары церков, которые могут иметь протоки абдоминальных желез [см. там же].
Однако если рассматривать эти признаки как синапоморфии, то трудно объяснить наличие синапоморфий у Symphyla с другими многоножками [см. ниже, Myriapoda (1) и Progoneata (1)–(4)].
III. Деление Atelocerata на Chilopoda и Labiata. Таксон Labiata Snodgrass 1938a {syn.circ.: Labiophora Kraus & Kraus 1993} объединяет Hexapoda, Symphyla и Collifera. Некоторые авторы считают его голофилетическим на основании следующих апоморфий.
(1) Считалось, что аутапоморфией Labiata является слияние пары максилл II в непарную нижнюю губу, или хилярий [см. ниже, Symphyla (4) и Hexapoda (3)]; однако в действительности у Collifera этот признак отсутствует, так как их хилярий образован не максиллами II, а максиллами I [см. ниже, Collifera (1) и (2)] (рис. 4.1).
(2) Наличие втяжных пузырьков, расположенных на коксах или оказавшихся на туловище в результате слияния коксы с туловищем [см. ниже, Progoneata (5), Symphyla (6), Diplopoda (8), Pauropoda (4) и Hexapoda (7)]. Однако подобные пузырьки описаны и для некоторых паукообразных и онихофор [ср.: Onychophora: «Классификация»: Peripatus/fg2; Chelicerata: «Классификации» III: Arachnida (5d)].
(3) То или иное отличие первых 3 туловищных сегментов (V–VII посторальных сегментов) от следующих: остальные туловищные сегменты у насекомых и личинок Collifera не имеют ног [см. ниже, Collifera (3) и Hexapoda (4)], а у взрослых диплопод несут по две пары ног на один тергит [см. ниже, Diplopoda (3)]; у симфил они чаще, чем первые три, имеют по два тергита на одну пару ног [см. ниже, Symphyla (5) и рис. 5.2.3].

IV. Деление Atelocerata на Amyocerata и Myocerata. Таксон Amyocerata объединяет Triplura и Pterygota на основании наличия мышц только в первом членике антенны и других апоморфий; он является голофилетическим и рассматривается ниже, как подчиненный таксон в составе Hexapoda. Таксон Myocerata Remington 1955 объединяет Myriapoda и Entognatha на основании симплезиоморфии – наличия мышц более чем в одном членике антенны (обычно во всех кроме последнего); этот таксон является парафилетическим и поэтому большинством авторов не признается.
V. Деление Atelocerata на Progoneata и Opisthogoneata. Таксон Progoneata характеризуется положением полового отверстия в передней части туловища впереди гонад и некоторыми другими апоморфиями. Он, по всей видимости, является голофилетическим и рассматривается ниже, как подчиненный таксон в составе Myriapoda.. Таксон Opisthogoneata Pocock 1893a, b {synn.circ.: Trignatha Lameere 1895 = Entomoptera Packard 1903; по циркумскрипции также соответствует «Insecta» sensu Kingsley 1894 (non Linnaeus 1758)} объединяет Chilopoda и Hexapoda лишь на основе симплезиоморфии – наличия полового отверстия в задней части туловища позади гонад.
VI. Деление Atelocerata на Myriapoda и Hexapoda. Здесь мы принимаем деление Atelocerata на Myriapoda и Hexapoda; каждый из этих таксонов является голофилетическим, что доказывается уникальными аутапоморфиями (см. ниже, главы V и VI).
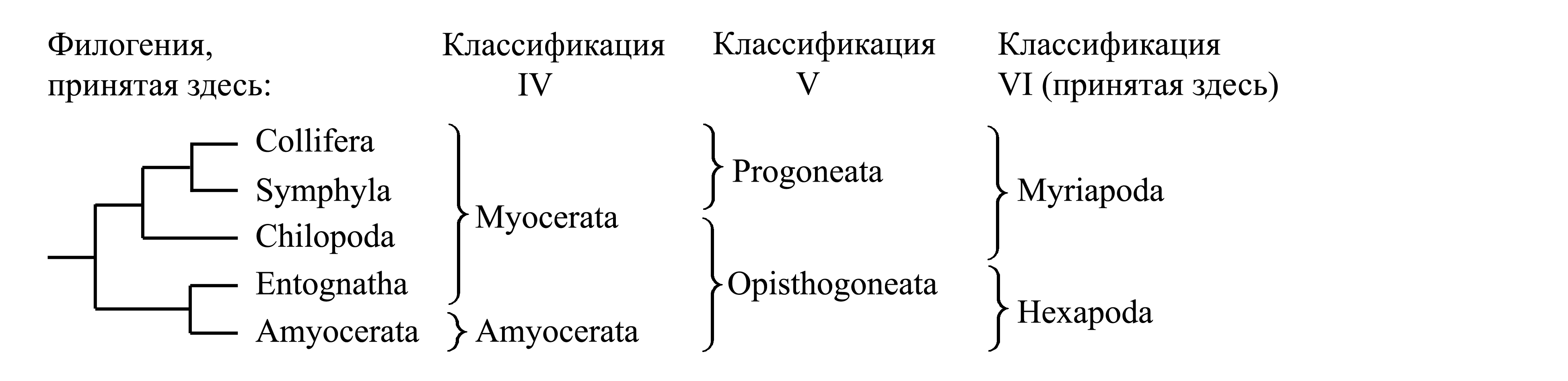
Здесь мы принимаем деление Atelocerata на два подчиненных таксона – Myriapoda (см. главу V) и Hexapoda (Глава VI, с. 0000). Кроме того, известны вымершие членистоногие, предположительно относимые к Myriapoda (см. главу V, с. 0000)
Atelocerata, или Pediculus/f2=Scarabaeus/g2
![]() 1. Myriapoda, или Scolopendra/fg1
1. Myriapoda, или Scolopendra/fg1
![]() 1.1. Progoneata, или Julus/fg1
1.1. Progoneata, или Julus/fg1
![]() 1.1.1. Symphyla, или Scolopendrella/fg1
1.1.1. Symphyla, или Scolopendrella/fg1
![]() 1.1.2. Collifera, или Julus/fg2
1.1.2. Collifera, или Julus/fg2
![]() 1.1.2.1. Diplopoda,
или Julus/fg3
1.1.2.1. Diplopoda,
или Julus/fg3
![]() 1.1.2.2. Pauropoda,
или Pauropus/fg1
1.1.2.2. Pauropoda,
или Pauropus/fg1
![]() 1.2. Chilopoda,
или Scolopendra/fg2
1.2. Chilopoda,
или Scolopendra/fg2
![]() 1.2.1. Notostigmophora,
или Scutigera/fg1
1.2.1. Notostigmophora,
или Scutigera/fg1
![]() 1.2.2. Pleurostigmophora,
или Scolopendra/fg3
1.2.2. Pleurostigmophora,
или Scolopendra/fg3
![]() Вымершие группы, предположительно относимые к Myriapoda
Вымершие группы, предположительно относимые к Myriapoda
![]() 2. Hexapoda, или Pediculus/f3=Scarabaeus/g3
2. Hexapoda, или Pediculus/f3=Scarabaeus/g3
![]() 2.1. Entognatha, или Podura/fg1
2.1. Entognatha, или Podura/fg1
![]() 2.2. Amyocerata, или Pediculus/f4=Scarabaeus/fg4
2.2. Amyocerata, или Pediculus/f4=Scarabaeus/fg4