КЛАДОЭНДЕЗИС НАСЕКОМЫХ

![]()
|
КЛАДОЭНДЕЗИС НАСЕКОМЫХ |
|||||||
|
|
|
fg | |||||
V-1.2. Chilopoda Latreille 1817 – хилоподы, или губоногие
(рис. 5.5.1–5.5.4)
(рис. 5.5.1, 5.5.2,
5.5.3, 5.5.4)
|
Названия, однозначно соответствующие по циркумскрипции: — Syngnatha Latreille 1802; — Chilopoda Latreille 1817; — Chilopodea Pearce 1936. По циркумскрипции также соответствует таксонам под типифицированными названиями, образованными от родового названия Scolopendra: Scolopendria, Scolopendrides и др. Иерархическое название: Scolopendra/f1=g2 [f:1814(fam.21); g:1758] (incl. Scutigera). |
|
Ранг. В XX веке – обычно класс или подкласс; в XIX веке – часто семейство или отряд.
Название. Общепринятым название этого таксона является Chilopoda, несмотря на то, что есть более старое название Syngnatha.
Аутапоморфии Chilopoda
(1) Строение мандибул. Мандибулы модифицированы (рис. 5.5.1A, 5.5.2C–D): премандибула и стипес [исходно сочлененные подвижно – см. Myriapoda (1)] слились неподвижно; задний конец стипеса погружен в карман и сочленен с головной капсулой не прямо, а посредством склерита, идущего по дну кармана; в результате этого вместо исходных четырех сочленений (головная капсула – фультура, фультура – премандибула, премандибула – стипеса, стипес – головная капсула) мандибула имеет четыре других сочленения: головная капсула – фультура, фультура – мандибула, мандибула – склерит кармана, склерит кармана – головная капсула).
Мандибула имеет косые швы, разделяющие ее на несколько склеритов, но при этом премандибула и стипес соединены неподвижно; несмотря на это, из премандибулы в стипес идет внутримандибулярная мышца (рис. 5.5.2C). На дорсальной стороне премандибулы имеется мыщелок, образующий сустав с фультурой, которая, в свою очередь, подвижно причленяется к краю головной капсулы (как у прочих Myriapoda) (рис. 5.5.2C). Задний конец стипеса мандибулы погружен в неглубокий челюстной карман, образованный налегающим снизу основанием хилярия [см. (2)] и подвижно соединен с краем головной капсулы посредством палочкообразного склерита, идущего по дну челюстного кармана. Так что мандибула и спереди, и сзади причленена к склеротизованной головной капсуле не прямо, а опосредованно, через подвижные склериты, лежащие на нижней мембранозной стороне головы; поэтому, несмотря на неподвижное соединение премандибулы со стипесом, мандибула может совершать сложные движения.
(2) Конечности III сегмента. Конечности III посторальной пары (следующей за мандибулами) и поэтому часто называемые «максиллами первыми», преобразованы в аналог нижней губы – хилярий (chilarium) (рис. 5.5.1A, 5.5.2D). Коксоподиты этих придатков сомкнуты между собой непосредственно позади гипофаринкса и отчасти срастаются с ним; телоподиты этих придатков 2-члениковые, недлинные, уплощенные и расширенные. В отличие от хилярия Collifera (который развит на том же сегменте), у Chilopoda отсутствует непарный вырост стернита. В отличие от максилл Symphyla и Hexapoda (развитых на том же сегменте), конечности, образующие хилярий у Chilopoda, неподвижные, не имеют гнатобаз на коксоподитах и не служат челюстями.
(3) Конечности IV сегмента. Конечности IV посторальной пары, следующие за хилярием и поэтому часто называемые «максиллами вторыми», состоят из плоских коксоподитов (у Pleurostigmophora сросшихся по средней линии) и расставленных телоподитов – щупиков (palpi). Щупик по строению сопоставим с телоподитом туловищной ноги того же животного [см. (7)], отличаются от нее лишь меньшими размерами и некоторыми упрощениями: у всех Notostigmophora редуцирован коготок, но сохраняются все прочие членики [см. ниже, Notostigmophora (2) и рис. 5.5.1A]; у всех Pleurostigmophora трохантер, фемур и пателла слиты, но сохраняется коготок [см. ниже, Pleurostigmophora (2) и рис. 5.5.2B]. Таким образом, у ближайшего общего предка Chilopoda щупики по строению ничем не отличались от туловищных ног, но утратили локомоторную функцию из-за того, что позади них расположены направленные вперед хватательные ногочелюсти [см. (4)]. Несмотря на различие в размерах и функции щупиков и туловищных ног, в строении этих конечностей сохраняется эволюционная корреляция. Вероятно, у Chilopoda конечности IV посторальной пары модифицировались в качестве ротовых независимо от других членистоногих, и сохранение явного сходства этих конечностей с туловищными ногами является плезиоморфией Chilopoda, уникальной среди Atelocerata.
Сегмент, несущий щупики, не имеет собственного тергита и, по-видимому, входит в состав цельной головной капсулы (вместе с I–III посторальными сегментами); в этом отношении Chilopoda сходны с Symphyla, Hexapoda и многими Eucrustacea, но отличаются от Collifera, у которых этот сегмент образует коллюм.
У зародыша эмбриональная кутикула щупика несет яйцевой зуб, служащий для прорывания яйцевой оболочки. Эту особенность рассматривают как аутапоморфию Chilopoda (Dohle 1985).
(4) Конечности V сегмента. Следующая за щупиками пара придатков (т.е. конечности V посторальной пары) преобразована в мощные хватательные ногочелюсти (maxillipedes) (рис. 5.5.1A, 5.5.2B). Коксоподиты ногочелюстей соприкасаются или слиты, образуя склеротизованную пластинку, прикрывающую снизу голову и ротовые органы. Телоподиты ногочелюстей по строению сопоставимы с телоподитами ног и щупиками [см. (7)]; трохантерофемур (слитые 2-й и 3-й членики) очень массивный; лапка и коготок, слитые вместе, образуют сильно склеротизованный хватательный крюк (ta+pta). Внутри ногочелюсти проходит проток ядовитой железы, открывающийся на вершине коготка. Уникальная аутапоморфия.
Сегмент, несущий ногочелюсти, в отличие от предыдущих, не входит в состав головной капсулы, и его тергит либо обособлен (у большинства Chilopoda), либо слит со следующим туловищным тергитом (у Oligostigmata) (рис. 5.5.4).
Chilopoda являются хищниками и используют ядовитые крючья для ловли и умерщвления добычи.
(5) Сегменты туловища. Сегменты туловища образуют диплосегменты [см. Myriapoda (3)] и один триплосегмент: позади трех диплосегментов следует триплосегмент, позади него еще три диплосегмента; позади них могут находиться дополнительные диплосегменты (у Chilopoda-Epimorpha).
Судя по всему, исходным для Chilopoda является следующее строение туловища (рис. 5.5.3). После ногочелюстного сегмента [см. (4)] имеется 15 сегментов, каждый из которых несет одну пару развитых ног; из них у I–VII сегментов тергит нечетного сегмента длинный, четного – короткий; тергиты VII и VIII сегментов длинные, и далее тергит каждого четного сегмента длинный, нечетного – короткий. У личинки первого возраста имеются только I–VII из этих сегментов, следующие сегменты (начиная с внеочередного длинного) добавляются в ходе постэмбрионального развития [т.е. имеется анаморфоз – см. Metameria (1b)]. Трахеостии [см. ниже, Notostigmophora (5) и Pleurostigmophora (7)] имеются только на сегментах с длинными тергитами (I, III, V, VIII, X, XII, XIV), причем на VII сегменте (длинном, за которым следует еще один длинный) трахеостии отсутствуют.
У разных Chilopoda эта схема строения видоизменена (рис. 5.5.4). Число сегментов, равное 15 и анаморфоз сохраняются у Notostigmophora, Ungulipalpi и Craterostigmophora (иногда объединяемых в таксон Anamorpha); при этом число пар ног у личинки первого возраста в этих группах варьирует: 4–7 пар у Notostigmophora, 6–8 (обычно 7) пар у Ungulipalpi, 12 пар у Craterostigmophora. Высказано предположение, что исходным числом пар ног у личинки первого возраста является не 7, а 4; это предположение основано на том, что у Notostigmophora обычно личинка вылупляется с 4 парами ног (рис. 5.5.1B), а в эмбриогенезе Lithobius forficatus зачатки первых 4 пар ног закладываются раньше других (Dohle 1985).
У Epimorpha число сегментов и ног не меняется в постэмбриогенезе (т.е. имеется эпиморфоз); при этом число сегментов увеличено по сравнению с исходным: имеется не менее 21 пары ног, причем у Pantastigmata число пар ног может быть неопределенным (т.е. варьирует у особей одного вида) и достигает 191. У всех Chilopoda число пар ног нечетное (Minelli & Bortoletto 1990); это связано с тем, что всегда, хотя бы в скрытой форме, сегменты объединены в диплосегменты, а VII–IX сегменты образуют триплосегмент.
Чередование длинных и коротких тергитов может быть хорошо выражено (у Ungulipalpi, Craterostigmophora, Devonobius/fg и некоторых Oligostigmata), либо короткие тергиты не отличаются от длинных и туловище выглядит гомономным (у Pantastigmata и некоторых Oligostigmata), либо, наоборот, короткие тергиты совсем исчезают и каждые два сегмента имеют единый тергит (у Notostigmophora – рис. 5.5.1A). В то время как у Notostigmophora число тергитов меньше числа пар ног, у Craterostigmophora, наоборот, некоторые длинные тергиты разделены на два, так что тергитов больше, чем пар ног.
Трахеостии могут быть редуцированы на некоторых сегментах (например, у отдельных представителей Ungulipalpi число пар трахеостий сокращается от исходного 7 до 2), а у Pantastigmata, наоборот, трахеостии развиты не через сегмент, а на всех сегментах.
(6) Генитальные сегменты. Позади последнего из туловищных сегментов, несущих ноги [т.е. пятнадцатого или более заднего – см. (5)], и впереди тельсона [см. Metameria (1b)] имеются два уменьшенных сегмента – прегенитальный и генитальный; эти сегменты у самца и/или у самки могут нести специализированные половые конечности – гоноподы (gonopodia) (рис. 5.5.3). Обе пары гонопод развиты только у самцов Notostigmophora, в остальных случаях пара гонопод развита лишь на генитальном сегменте. Если гоноподы хорошо развиты, прослеживается гомология их члеников с члениками ноги [см. (7)]. Гоноподы не служат для внутреннего оплодотворения; оплодотворение внешнее сперматофорное.
У самца и у самки половое отверстие непарное, открывается на генитальном сегменте, т.е. на конце тела позади гонад (в отличие от Progoneata); гонодукты у обоих полов парные; гонады самки (яичники) слиты в непарное образование; гонады самца (семенники) обычно парные, лишь у Ungulipalpi непарные (см. ниже, Pleurostigmophora: «Классификация»).
Признаки Chilopoda неясного филогенетического значения
(7) Строение ног. Нога состоит из 6 члеников, не считая претарсус (рис. 5.5.2B); их гомология с члениками ног насекомых вызывает разногласия (рис. 4.8). Кокса (1-й членик) всегда прикреплена к туловищу латерально. Трохантер (2-й членик) короткий, иногда слит с фемуром (3-м члеником). Лишь сочленение коксы и трохантера двухмыщелковое, тогда как все следующие сочленения одномыщелковые. Тарсус (6-й членик) бывает не расчленен, либо разделен на 2 или 3 вторичных членика, либо разделен на множество вторичных члеников (у некоторых Ungulipalpi и у Notostigmophora – рис. 5.5.1A). Претарсус [см. Atelocerata (3)] имеет вид непарного коготка.
Ноги последней пары волочащиеся, длиннее других и направлены назад, а у Pantastigmata и некоторых Oligostigmata они лишены коготков и имеют внешнее сходство с церками насекомых, в связи с чем их иногда называют «церками» [ср.: Hexapoda (8a)].
Размеры. Средние или крупные – 10–260 мм.
Геологический возраст. Девон – ныне (рис. 2.2) (см. ниже, Pleurostigmophora: «Классификация»: Devonobius/fg). Есть менее достоверные указания на нахождение Chilopoda в силуре (см. ниже, Notostigmophora).
Систематическое положение Chilopoda. Помимо принятого здесь отнесения Chilopoda непосредственно к Myriapoda, предлагались следующие классификации.
I. Объединение Chilopoda, Symphyla и Hexapoda в таксон Etymachila (см. Atelocerata: «Классификации» I).
II. Объединение Chilopoda и Collifera в таксон Monomalata (см. Atelocerata: «Классификации» II).
III. Объединение Chilopoda и Hexapoda в таксон Opisthogoneata (см. Atelocerata: «Классификации» V).
IV. Объединение Chilopoda и Symphyla в таксон Homopoda (см. Myriapoda: «Классификации» I).
Классификации Chilopoda
I. Деление Chilopoda на Anamorpha и Epimorpha. Таксон Anamorpha Haase 1880 {syn.circ.: Ovodispersa Prunescu 2006} характеризуется тем, что число туловищных сегментов и пар ног у взрослых животных постоянно и равно 15, а развитие анаморфное, т.е. у личинки число туловищных сегментов меньше пятнадцати и увеличивается с возрастом; самка откладывает яйца порознь и не заботится о них. Таксон Epimorpha Haase 1880 характеризуется тем, что число сегментов и пар ног больше пятнадцати, а развитие эпиморфное, т.е. число сегментов не меняется с возрастом; самка откладывает яйца в одном месте и охраняет их. Эта классификация широко признана теми, у кого нет мнения о полярности данных признаков. В зависимости от разных представлений о полярности признаков, один из этих таксонов считают парафилетическим и расформировывают, а другой считают голофилетическим и сохраняют: либо расформировывают Epimorpha и сохраняют Anamorpha (см. II), либо расформировывают Anamorpha и сохраняют Epimorpha (см. III). Некоторые авторы определяют полярность признаков исходя из общих соображений о том, в каком направлении должна идти эволюция: по одной версии – от большого числа сегментов к малому, по другой – от малого к большому; по одной версии – от анаморфоза к эпиморфозу, по другой – от эпиморфоза к анаморофозу. Однако таких общих законов эволюции не существует (см. раздел I.4.2); в ходе эволюции различных таксонов членистоногих число сегментов многократно и уменьшалось, и увеличивалось; анаморфоз и эпиморфоз независимо возникали много раз. Правильный вывод о полярности признаков может быть сделан только при анализе уникальных аутапоморфий, а не таких повторяющихся признаков, как полимерность и олигомерность или анаморфоз и эпиморфоз. Анализ уникальных признаков хилопод приводит к выводу, что в данном случае Anamorpha является парафилетическим таксоном, а Epimorpha – голофилетическим (см. III).

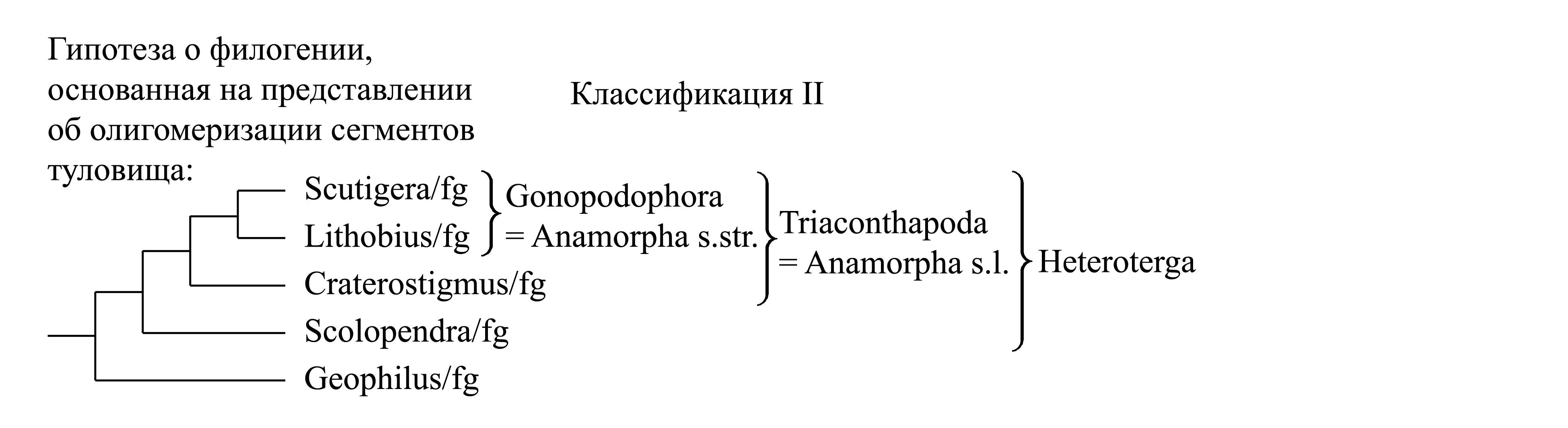
II. Деление Chilopoda на Pantastigmata и Heteroterga. Эта классификация основана на том, что гомономность сегментации туловища, большое неопределенное число пар ног и эпиморфоз считаются плезиоморфиями, а чередование длинных и коротких тергитов, постоянное число пар ног и анаморфоз считаются апоморфиями (Ax 1999). Таксон Heteroterga Ax 1999 характеризуется чередованием длинных сегментов с дыхальцами и коротких сегментов без дыхалец (в отличие от Geophilomorpha). Таксон Triakontapoda Ax 1999 (= Anamorpha s.l.) характеризуется резким различием между длинными и короткими тергитами и 15 парами ходильных ног (в отличие от большего числа ног у Scolopendromorpha и Geophilomorpha). Таксон Gonopodophora Ax 1999 (= Anamorpha s.str.) характеризуется наличием гонопод и откладкой яиц поодиночке. Согласно этому представлению о филогении таксон Epimorpha считается парафилетическим и в классификации отсутствует. В данном случае вывод о полярности признаков делался на основе лишь общих соображений о том, что эволюция должна идти в сторону олигомеризации гомологичных органов; при этом уникальные аутапоморфии Epimorpha не приняты во внимание.
III. Деление Chilopoda на Notostigmophora и Pleurostigmophora. Эта классификация основана на иной гипотезе о филогении. Каждая ветвь этой филогенетической схемы обосновывается уникальными аутапоморфиями (Dohle 1985, Shear & Bonamo 1988) [см. ниже, Notostigmophora (1)–(5), Pleurostigmophora (1)–(7) и «Классификация Pleurostigmophora»]. На основе такого представления о филогении хилопод делается вывод, что у Epimorpha гомономность сегментации и большое неопределенное число сегментов туловища являются не примитивными чертами, а апоморфиями.
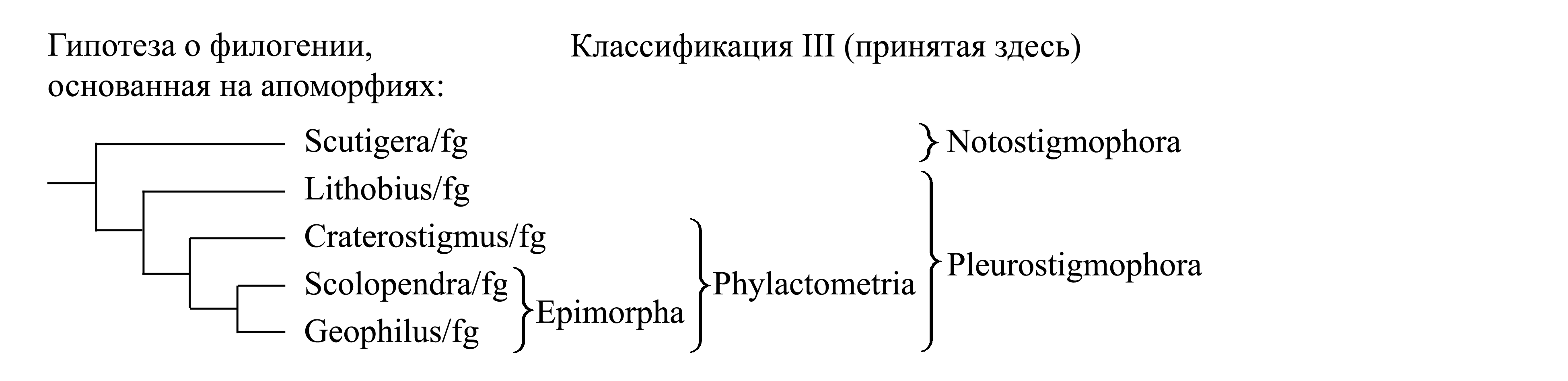
Здесь мы принимаем деление Chilopoda на два подчиненных таксона – Notostigmophora и Pleurostigmophora (стр. 0000).
Chilopoda, или Scolopendra/fg2
![]() 1. Notostigmophora,
или Scutigera/fg1
1. Notostigmophora,
или Scutigera/fg1
![]() 2. Pleurostigmophora,
или Scolopendra/fg3
2. Pleurostigmophora,
или Scolopendra/fg3