КЛАДОЭНДЕЗИС НАСЕКОМЫХ

![]()
|
КЛАДОЭНДЕЗИС НАСЕКОМЫХ |
|||||||
|
|
|
fg | |||||
Глава V. Систематика Myriapoda
V-1. Myriapoda Latreille 1802 – многоножки
(рис. 5.1–5.5.4)
(рис. 5.1 ; 5.2.1,
5.2.2, 5.2.3 ; 5.3
; 5.4.1, 5.4.2
; 5.5.1,
5.5.2,
5.5.3, 5.5.4)
|
Названия, однозначно соответствующие по циркумскрипции: — Polypoda Lamarck 1801 (исходно во французском написании Polypodes) (non Polypoda Batsch 1789, nec Polypoda Guilding 1826, nec Polypoda Brandt 1841); — Myriapoda Latreille 1802 (non Myriapodes Latreille 1796); — Millepeda Leach 1814; — Myriapodia Rafinesque 1815; — Stigmatopnea Goldfuss 1820; — Myriapodes tracheales Burmeister 1830. Иерархическое название: Julus/f1=Scolopendra/g1 [f:1814(fam.20); g:L.1758(gen.242)]. |
|
Ранг. В XX веке – обычно надкласс или класс; в XIX веке – от семейства до класса.
Название. Общепринятым названием этого таксона является Myriapoda. Исходно это название во французском написании Myriapodes Latreille 1796 относилось к большему таксону, включавшему не только многоножек, но и наземных ракообразных; в латинском написании название Myriapoda Latreille 1802 исходно относится к таксону, включающему только многоножек в современном понимании. Бурмеистер (Burmeister 1830) использовал оба названия как латинские: Myriapodes для того же таксона, что и Латрейль в 1796, и Myriapoda для подчиненного ему таксона того же объема, что Латрейль в 1802 (как синоним Myriapodes tracheales). Название Myriapoda можно употреблять в общепринятом значении, если следовать интерпретации Бурмеистера и считать название Myriapodes особым латинским названием, отличным от Myriapoda.
Аутапоморфии Myriapoda
(1) Модификация мандибул. Мандибулы [одночлениковые – см. Atelocerata (4)] имеют особое строение (рис. 5.1). Мандибула обычно разделена на переднюю выступающую жующую часть, называемую премандибула (praemandibula), или мандибулярная мала (mala mandibularis) (в англоязычной литературе – также «gnathal lobe») и заднюю невыступающую часть – стипес (stipes). У некоторых Diplopoda задняя часть, в свою очередь, подразделена швом на две подвижно сочлененные доли, называемые в этом случае стипес и кардо (cardo). У Symphyla и Diplopoda стипес мандибулы выглядит как часть боковой стенки головы, поэтому некоторые авторы ошибочно считали его «щекой», а «мандибулой» называли только премандибулу. Каждая из этих частей мандибулы (и премандибула, и стипес) имеет непосредственное сочленение с головой и мышцы, идущие из нее прямо в голову, так что эти части мандибулы являются не настоящими члениками конечности, а результатом вторичного расчленения коксоподита. Вполне подвижное сочленение премандибулы со стипесом имеется у Symphyla, Diplopoda и Pauropoda-Hexamerocerata, тогда как у Chilopoda сохраняется лишь шов, не обеспечивающий настоящей подвижности. От внутренней стенки стипеса к эндоскелету головы (либо к мезодермальному лигаменту, либо к эктодермальным апофизам) идут мышцы, вполне сходные по своему месту прикрепления с вентральными аддукторами мандибул других Mandibulata. От края премандибулы начинается аподема, от которой отходит мощная мышца, идущая к внутренней стенке головной капсулы, – это дорсальный аддуктор мандибулы (т.е. приводящая мышца, идущая к тергиту) (рис. 5.1B: ADm); он проходит спереди (при прогнатном положении головы – дорсальнее) мышц, идущих от стипеса, так что соответствует передней дорсальной мышце мандибул других Mandibulata (рис. 4.6A: ADm). Задней дорсальной мышцы (которая бы проходила позади или вентральнее мышц стипеса) у Myriapoda нет. У других Mandibulata – Eucrustacea и Hexapoda – наоборот: задняя дорсальная мышца мандибул (PDm) обычно мощная и служит аддуктором (приводящей мышцей), а передняя дорсальная мышца (ADm) слабее и служит абдуктором (отводящей мышцей) (рис. 4.6A) [см. Mandibulata (1)]. Так что по своему расчленению и мускулатуре мандибулы многоножек похожи не на мандибулы, а на максиллы насекомых [ср.: Hexapoda (2) и рис. 6.7.1D].
На вентральной мембранозной поверхности головы непосредственно перед мандибулами лежат парные подвесы гипофаринкса, или фультуры (fulturae) (рис. 5.1A: flt) – склериты, медиально прикрепленные к гипофаринксу; латеральные концы фультур могут быть подвижно сочленены с краями головной капсулы. От фультур в глубь головы отходит пара аподем (рис. 5.1A: ap); эти аподемы иногда считают гомологичными передними руками тенториума Amyocerata (Manton 1964); в отличие от тенториума они подвижны относительно головной капсулы. Премандибулы могут быть сочленены с подвижными фультурами, что позволяет им совершать сложные движения.
Авторы, придерживавшиеся теории о полифилетическом происхождении членистоногих, считали расчлененность мандибул у Myriapoda плезиоморфией, поскольку считали их мандибулы произошедшими не из проксимального членика (базиподита) исходной конечности членистоногих, а из целой конечности онихофор [см. Mandibulata (1)]. Премандибула и стипес мандибулы многоножек по строению и мускулатуре напоминают лацинию и стипес максиллы насекомых [ср.: Hexapoda (2)]; поскольку максилла насекомых несет членистый щупик (телоподит), нет сомнений в том, что ее стипес и лациния являются производными одного членика – коксоподита. Судя по всему, то же относится и к подвижно сочлененным склеритам мандибулы многоножек.
В отличие от насекомых, у которых следующая за мандибулами пара конечностей представляет собой максиллы с подвижно расчлененными коксоподитами [ср.: Hexapoda (2)], у большинства многоножек таких придатков нет: у диплопод, пауропод и хилопод за мандибулами следуют неподвижные конечности, образующие хилярий и выполняющие функцию нижней губы [см. ниже, Collifera (1) и Chilopoda (2)]; лишь у симфил имеются максиллы, но более простого строения, чем у насекомых [см. ниже, Symphyla (3)]. В связи с отсутствием подвижных максилл, мандибулы многоножек должны выполнять сложные движения, манипулировать частицами пищи и отправлять их в рот. Благодаря тому, что мандибула соединена с головной капсулой не двумя, а четырьмя подвижными сочленениями (головная капсула – фультура, фультура – премандибула, премандибула – стипес, стипес – головная капсула), она способна совершать такие движения. У хилопод утрачена подвижность в соединении премандибулы со стипесом, но вместо сочленения стипеса с головной капсулой появились два новых сочленения, так что общее число сочленений также равно четырем [см. ниже, Chilopoda (1)].
(2) Утрата глазков. Из двух зрительных систем [см. Euarthropoda (4)], простые глазки всегда отсутствуют, а фасеточные глаза имеют тенденцию к редукции. Типичное строение кристаллического конуса [см. Mandibulata (4)] утрачено: лишь у Chilopoda-Notostigmophora имеются настоящие фасеточные глаза с особым строением кристаллического конуса (рис. 5.5.1C–D), у всех остальных Myriapoda глаза распадаются на отдельные простые глазки, не имеющие кристаллического конуса. Неясно, является ли наличие фасеточных глаз у Notostigmophora исходным (т.е. плезиоморфией, уникальной среди Myriapoda), или новообразованием (т.е. аутапоморфией) [см. ниже, Notostigmophora (6)]. У всех Symphyla, всех Pauropoda, некоторых Diplopoda и некоторых Pleurostigmophora глаза утрачены [см. ниже, Symphyla (1) и Pauropoda (1)]. Редукция глазков и глаз – неуникальная апоморфия, связанная с подстилочным и почвенным образом жизни многоножек; то же у многих Hexapoda – у всех Entognatha и некоторых Amyocerata.
Признаки Myriapoda неясного филогенетического значения
(3) Диплосегментность. Имеется тенденция к объединению туловищных сегментов по два; такие повторяющиеся отделы, состоящие их двух сегментов каждый, называются диплосегменты. Диплосегментность явно выражена у всех Diplopoda, многих Pauropoda и Chilopoda; в скрытой форме она имеется у всех Myriapoda, включая Symphyla [см. ниже, Symphyla (5), Diplopoda (3), Pauropoda (4) и Chilopoda (5)]. Диплосегмент может иметь два разных тергита и две пары ног (у большинства Chilopoda-Pleurostigmophora), либо единый слитный тергит и две пары ног (например, у Chilopoda-Notostigmophora и на брюшке у Diplopoda), либо два тергита и одну пару ног (у Chilopoda-Craterostigmophora и отчасти у Symphyla), либо единый тергит и одну пару ног (такое состояние достоверно встречается у последних развивающихся сегментов брюшка Diplopoda и предполагается для груди Diplopoda). В последнем случае невозможно отличить диплосегмент от гаплосегмента.
У других членистоногих попарное объединение сегментов не наблюдается: у них если сегменты объединяются в комплексы, то такие комплексы (тагмы) в теле животного не повторяются. Помимо Myriapoda явно выраженные диплосегменты имеются только у вымершего таксона Euthycarcinus/fg1 (см. ниже, Вымершие группы). Однако возможно, что явление диплосегментности не является апоморфией Myriapoda, а может быть объяснено механизмом морфогенеза (Minelli & Bortoletto 1988, 1990): у некоторых насекомых на стадии эмбриогенеза, предшествующей сегментации, обнаружены белки, которые маркируют сразу по два будущих сегмента (Akam 1994, Patel & al. 1994). Кроме того, существует предположение о наличии у насекомых диплосегментов, утративших вторую пару ног и потому неотличимых от гаплосегментов [ср.: Hexapoda (4c)].
(4) Гомономность туловища. Туловище не подразделено на грудь и брюшко в том смысле, в каком эти термины употребляются применительно к Hexapoda и Eucrustacea [см. Metameria (1c)]: у многоножек все сегменты туловища либо несут по паре ног, либо пары ног каким-либо иным образом равномерно распределены по всей длине туловища [см. (3) и ниже, Symphyla (5), Diplopoda (3), Pauropoda (4) и Chilopoda (5)]. Все туловищные сегменты и ноги всех пар сходны, и в их строении хорошо выражена эволюционная корреляция.
Обычно гомономную сегментацию туловища Myriapoda считают плезиоморфией, однако высказана противоположная точка зрения о том, что это апоморфия (Щербаков 1996a). Основанием для этого служит тот факт, что разделение туловища на две тагмы – грудь (куда входит то или иное число сегментов с локомоторными конечностями) и брюшко (куда входит то или иное число сегментов с недоразвитыми конечностями) наблюдается у разных членистоногих, в частности у Eucrustacea и Hexapoda; при этом есть данные о том, что у различных членистоногих, включая изученных представителей Eucrustacea и Hexapoda, эта дифференциация регулируется гомологичными гомеотическими генами (Carroll 1995). Отсюда можно сделать вывод, что гетерономное строение туловища первично для Mandibulata, а гомономность появилось вторично у Myriapoda (а также независимо от них у Remipedia среди Eucrustacea).
Плезиоморфии и изменчивые признаки Myriapoda
(5) Состав головы. Строение ротовых конечностей, находящихся позади мандибул [см. (1)], различно у разных Myriapoda (рис. 4.1): у симфил имеется пара максилл и непарный нижняя губа [см. ниже, Symphyla (3) и (4)]; у диплопод и пауропод лишь непарный хилярий [см. ниже, Collifera (1)]; у губоногих имеются хилярий, ногоподобные щупики и специализированные ядовитые ногочелюсти [см. ниже, Chilopoda (2)–(4)]. В связи с этим возникли принципиальные разногласия об исходном сегментарном составе головы Myriapoda. По одной версии исходно в ее состав входит 4 посторальных сегмента, включая IV сегмент, несущий максиллы II; в этом случае по строению головы Myriapoda принципиально сходны с Hexapoda; предполагалось, что такой же сегментарный состав головы имеется у всех Eucrustacea и исходно свойственен Mandibulata (Snodgrass 1935, 1938). По другой версии в составе головы Myriapoda изначально было лишь 3 посторальных сегмента, а IV сегмент вошел в состав головы независимо у разных Myriapoda; об этом говорит тот факт, что у Collifera четвертый сегмент является туловищным, хотя и потерявшем конечности [см. ниже, Collifera (2)], а у Chilopoda он вошел в состав головы, но его конечности (щупики) сохранили строение, свойственное туловищным ногам [см. ниже, Chilopoda (3)]. В прошлом на основании этого делали вывод, что Diplopoda не имеют отношения к таксону, объединяющему Eucrustacea и Hexapoda. Однако после открытия Cephalocarida Sanders 1955 стало известно, что у Eucrustacea конечности четвертого посторального сегмента также исходно не специализированы в качестве ротовых [см. Mandibulata (2)].
Геологический возраст. Палеозой – ныне. О древнейших представителях см. ниже, Diplopoda.
Систематическое положение Myriapoda. Помимо принятого здесь отнесения Myriapoda к Atelocerata, предлагались следующие классификации.
I. Отнесение Myriapoda непосредственно к Mandibulata (см. Mandibulata: «Классификации» II).
II. Объединение Myriapoda и Entognatha в парафилетический таксон Myocerata (см. Atelocerata: «Классификации» VI).
III. Объединение Myriapoda и Chelicerophora в таксон Myriochelata Pisani & al. 2004 {syn.circ.: Paradoxopoda Mallatt & al. 2004}. Этот таксон не имеет никакого диагноза и был установлен лишь путем некорректных вычислений на основе принципа парсимонии (см. I.4.2.6).
Статус Myriapoda. Существует распространенное мнение, что Myriapoda – парафилетический таксон, предковый для Hexapoda и отличающийся от Hexapoda лишь плезиоморфиями; в частности, плезиоморфией Myriapoda считают большое число пар одинаковых туловищных конечностей, хотя на этот счет имеется и противоположное мнение [см. (4)]. По мнению некоторых авторов, одна из групп в составе многоножек (либо Symphyla, либо Chilopoda) является сестринской для Hexapoda и может быть объединена с ними в один таксон. В соответствии с этим в некоторых классификациях таксон Myriapoda отсутствует (см. Atelocerata: «Классификации» I, II, III и V). Некоторые авторы даже считают многоножек, а вместе с ними и Atelocerata, полифилетической группой (Старобогатов 1991 и др.). Представления о парафилии или о полифилии многоножек в значительной мере связаны с большим разнообразием их ротовых придатков, что указывает на независимое формирование головного отдела у разных многоножек [см. Myriapoda (5) и ниже, Symphyla (3), (4), (10), Collifera (1), (2) и Chilopoda (2), (3)].
Однако существует мнение о том, что Myriapoda – это голофилетический таксон (Dohle 1980 и др.), что обосновывается приведенными выше апоморфиями.
Классификации Myriapoda. К Myriapoda относятся 4 четко очерченных таксона – Symphyla, Diplopoda, Pauropoda и Chilopoda (см. ниже).
В литературе иногда упоминается о существовании описанного из Индии вида Polyxenella krishnani Rajulu 1963, помещавшегося в состав Symphyla, но отличающегося от всех Symphyla задним положением полового отверстия и другими признаками, в корне меняющими сложившиеся представления о системе многоножек. В действительности здесь речь идет о неквалифицированно описанном представителе Diplopoda-Penicillata; в его описании половое отверстие перепутано с анальным и допущены другие грубые ошибки (Dohle 1980). Все современные виды многоножек бесспорно распределяются по названным четырем таксонам, лишь некоторые ископаемые формы имеют спорное положение из-за недостаточной изученности.
Некоторые авторы рассматривали в качестве особого таксона того же ранга, что Diplopoda и Chilopoda, вымерший карбоновый таксон Archipolypoda Scudder 1882 {synn.circ.: Macrosterni Fritsch 1899 = Palaeocoxopleura Verhoeff 1928; nom.typ.: Euphoberia/fg [f:1882; g:1868]}. Это было связано с ошибочным представлением об общей характеристике диплопод; сейчас Archipolypoda помещаются в состав Diplopoda. Архиполиподы имеют такое же строение диплосегментов брюшка и трех гаплосегментов груди, что и прочие диплоподы [см. ниже, Diplopoda (3)] (Burke 1979). Однако в отличие от некоторых современных диплопод, имеющих втяжные пузырьки на коксах, у Archipolypoda по паре втяжных пузырьков имеется на каждом стерните между коксами каждой пары ног, подобно Symphyla (Hannibal 1995, 1998).
О некоторых животных неясного систематического положения, лишь предположительно относимых к Myriapoda, см. ниже.
Существуют разногласия по поводу того, как следует группировать таксоны Symphyla, Diplopoda, Pauropoda и Chilopoda в таксоны более высоких рангов; предлагались следующие классификации.
I. Деление Myriapoda на Collifera и Homopoda. Таксон Collifera является голофилетическим и характеризуется набором аутапоморфий; мы рассматриваем его как подчиненный таксон в составе Progoneata – см. ниже (с. 000).
Таксон Homopoda Pocock 1893a {syn.circ.: Atelopoda Boudreaux 1979a; по циркумскрипции также соответствует Chilopoda в широком понимании, принятом в середине XIX века}. Объединяет Symphyla и Chilopoda. Помимо явных плезиоморфий, для Homopoda указывался следующий признак (Boudreaux 1979a): IV посторальный сегмент входит в состав головы; этот сегмент у симфил несет слитную нижнюю губу [см. ниже, Symphyla (4)], а у Chilopoda он несет пару ногоподобных придатков [см. ниже, Chilopoda (3)]. Помимо Homopoda, этот же сегмент входит в состав головы у Hexapoda и многих Eucrustacea (рис. 4.1), так что данный признак не может служить доказательством голофилии Homopoda.
II. Деление Myriapoda на Progoneata и Chilopoda. Каждый из этих таксонов является голофилетическим и имеет аутапоморфии – см. ниже, характеристики Progoneata и Chilopoda (с. 000).
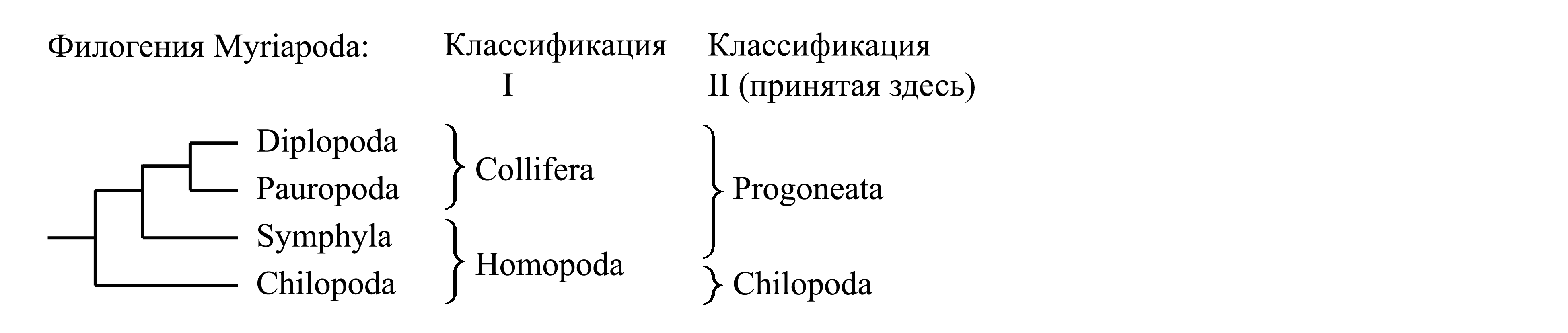
Здесь мы принимаем деление Myriapoda на два подчиненных таксона – Progoneata и Chilopoda (стр. 0000).
Myriapoda, или Scolopendra/fg1
![]() 1.
Progoneata, или Julus/fg1
1.
Progoneata, или Julus/fg1
![]() 1.1. Symphyla, или
Scolopendrella/fg1
1.1. Symphyla, или
Scolopendrella/fg1
![]() 1.2. Collifera, или Julus/fg2
1.2. Collifera, или Julus/fg2
![]() 1.2.1. Diplopoda, или Julus/fg3
1.2.1. Diplopoda, или Julus/fg3
![]() 1.2.2. Pauropoda, или
Pauropus/fg1
1.2.2. Pauropoda, или
Pauropus/fg1
![]() 2. Chilopoda, или Scolopendra/fg2
2. Chilopoda, или Scolopendra/fg2
![]() 2.1. Notostigmophora, или
Scutigera/fg1
2.1. Notostigmophora, или
Scutigera/fg1
![]() 2.2. Pleurostigmophora, или
Scolopendra/fg3
2.2. Pleurostigmophora, или
Scolopendra/fg3
![]() Вымершие группы, предположительно
относимые к Myriapoda
Вымершие группы, предположительно
относимые к Myriapoda