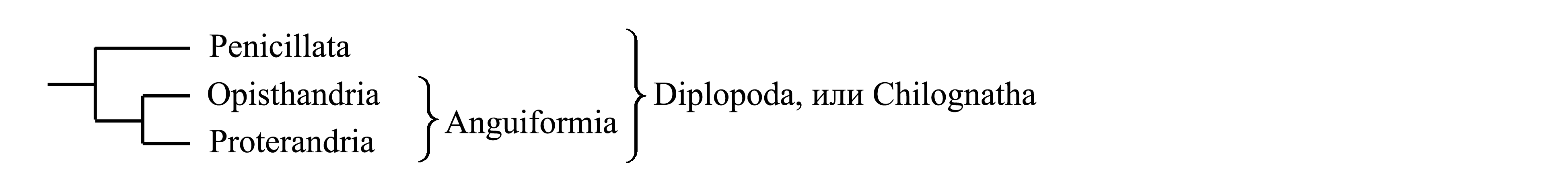КЛАДОЭНДЕЗИС НАСЕКОМЫХ

![]()
|
КЛАДОЭНДЕЗИС НАСЕКОМЫХ |
|||||||
|
|
|
fg | |||||
V-1.1.2.1. Diplopoda Blainville in Gervais 1844 – диплоподы, или двупарноногие
(рис. 5.3)
|
Названия, соответствующие по циркумскрипции: — Chilognatha Latreille 1802; — Diplopoda Blainville in Gervais 1844; — Diplopodea Pearce 1936. Иерархическое название: Julus/f4=g3 [f:1814; g:1758] (sine Pauropus; incl. Polyxenus). |
|
Ранг. В XX веке – обычно класс или подкласс; в XVIII–XIX веках – рода до класса.
Название. Общепринятым названием этого таксона в настоящее время является Diplopoda, несмотря на то, что название Chilognatha значительно старше и в свое время широко употреблялось. Строго говоря, оба названия являются неоднозначными, так как они были впервые опубликованы тогда, когда ближайший таксон Pauropoda не был известен. В работе Лаббока (Lubbock 1868), в которой впервые упоминается таксон Pauropoda, название Diplopoda было использовано для таксона, не включающего Pauropoda, и в таком узком значении название Diplopoda общепринято. Однако некоторые авторы использовали название Diplopoda в широком значении, т.е. для таксона, включающего Pauropoda (Kenyon 1895).
Аутапоморфии Diplopoda
(1) Строение антенн. У антенны [т.е. антенны I – см. Atelocerata (1)] последний членик на вершине несет 4 сложные щетинкоподобные сенсиллы конической формы [см. Euarthropoda (5c)]; иногда число этих сенсилл вторично увеличено.
Антенны всегда 8-члениковые (рис. 5.3A, D), но один или два последних членика могут быть погружены внутрь предыдущих, тогда антенны кажутся семи- или шестичлениковыми. Все членики антенны кроме последнего с собственной мускулатурой. У личинки первого возраста антенны 5-члениковые (рис. 5.3C), при последующих линьках число члеников увеличивается до восьми за счет развития проксимальных члеников. Возможно, стабильное число члеников антенны также является аутапоморфией Diplopoda; но возможно, что это плезиоморфия по отношению к Pauropoda, у которых число члеников уменьшено.
(2) Строение мандибул. Мандибулы [единственные подвижные конечности ротового аппарата – см. Collifera (1)] адаптированы к перетиранию растительных тканей: режущий край премандибулы образован стопкой зазубренных пластинок (которые иногда, например у Penicillata, сливаются вместе) и проксимальным цельным склеротизованным выступом, называемым молой (неясно, гомологичен ли этот выступ моле Eucrustacea и Hexapoda).
Сочленение премандибулы и стипеса подвижное [см. Myriapoda (1)]; при этом стипес мандибулы образует боковую стенку головы, в связи с чем его иногда ошибочно называют «щекой» или «головным плейритом»; стипес исходно цельный (рис. 5.3C–D), но иногда он в свою очередь расчленен на две подвижно соединенные части, называемые в этом случае стипес и кардо (рис. 5.3A). Аподемы фультуры короткие, так что мандибулярные мышцы крепятся не к ним, а к мезодермальному лигаменту.
У Colobognatha ротовой аппарат сосущий, с сильно видоизмененными колющими мандибулами.
(3) Сегменты туловища. Сегменты туловища, будучи однообразными [см. Myriapoda (4)], дифференцированы следующим образом: первые три пары ног, имеющиеся начиная с первого возраста [см. Collifera (3)] расположены по одной паре на трех первых сегментах, следующих после коллюма [см. Collifera (2)], а остальные ноги, добавляющиеся в постэмбриогенезе, расположены по две пары на следующих сегментах. Отдел туловища, состоящий из трех первых сегментов с тремя парами ног принято называть грудь (thorax), а следующий за ним отдел туловища принято называть брюшко (abdomen). Здесь термины «грудь» и «брюшко» употребляются в ином смысле, нежели применительно к насекомым и ракообразным: если у насекомых и ракообразных грудью называется часть туловища с развитыми локомоторными конечностями, а брюшком – часть туловища без них, то у диплопод отделы туловища, называемые грудью и брюшком, не различаются по строению и размеру находящихся на них конечностей; наоборот, у диплопод бóльшая часть ходильных конечностей находится на брюшке. Применительно к диплоподам термины «грудь» и «брюшко» связаны с предположением о гомологии между их трехсегментной грудью и трехсегментной грудью насекомых [см. рис. 4.1 и Collifera (3); ср.: Hexapoda (4)].
Туловище диплопод имеет следующее строение.
Коллюм, или шейный сегмент [первый, безногий сегмент туловища – см. Collifera (2)], имеет хорошо развитый тергит, который по форме отличается от других тергитов туловища; тергит коллюма бывает крупнее других (рис. 5.3A) или меньше других (рис. 5.3D). На вентральной стороне, непосредственно позади хилярия имеется пластинка, называемая гипостомом (hypostoma), или гулой (gula) (рис. 5.3A–B); предполагают, что гипостом является стернитом коллюма. Коллюм не имеет ни конечностей, ни их вестигиев, ни втяжных пузырьков (в отличие от Pauropoda), ни каких-либо иных парных придатков (см. ниже).
Следующие после коллюма три сегмента туловища образуют грудь; каждый из трех сегментов груди несет по одной паре ног [которые, в отличие от последующих пар ног, имеются у личинки начиная с первого возраста – см. Collifera (3)]. Иногда стерниты передних сегментов туловища смещены вперед, так что ноги кажутся отходящими от предыдущего сегмента; поэтому в составе груди некоторых диплопод ошибочно указывали не три, а четыре или пять пар ног (последние из которых в действительности относятся к брюшку); по этой же причине первая пара ног иногда кажется отходящей от коллюма, У половозрелых самцов некоторых Eugnatha ноги первой пары, будучи смещены вперед, сильно видоизменены, так что могут быть приняты за особые придатки коллюма (но у неполовозрелых особей они имеют такое же строение, как остальные ноги). В действительности же у всех Diplopoda коллюм не имеет никаких придатков, а следующие за ним три сегмента всегда имеют по одной паре конечностей.
Половые отверстия [парные, находящиеся позади второй пары ног – см. Collifera (4)], открываются на паре половых придатков: у самок этими придатками являются вульвы (vulvae), погруженные в полости стернитов, у самцов – это выдвигающиеся наружу пенисы (penes) (рис. 5.3A). У некоторых диплопод обособленных половых придатков нет, и гонопоры открываются на тазиках второй пары ног — это самцы Penicillata, Opisthandria и некоторых Proterandria и самки Penicillata и Opisthandria-Glomeridesmus/fg.
Брюшко состоит из явно выраженных диплосегментов [см. Myriapoda (3)]: каждый диплосегмент брюшка несет две пары ног, но имеет один цельный тергит (рис. 5.3A, D). Число диплосегментов брюшка, несущих по две пары ног, у разных диплопод бывает от 4 до 160; предполагают, что исходным является небольшое число сегментов, свойственное Penicillata и Opisthandria (см. ниже). Последние один или несколько диплосегментов брюшка не имеют ног.
У личинки первого возраста [имеющей только три пары ног, находящихся на трех сегментах груди – см. Collifera (3)] брюшко обычно имеет 1–4 (редко большее число) безногих диплосегментов и тельсон [см. (4)] (рис. 5.3C). При каждой последующей линьке тело удлиняется на один или несколько диплосегментов, добавляется по 1, по 2 либо по большему числу пар ног; так что на некоторых стадиях развития позади диплосегментов с двумя парами ног может быть один или несколько диплосегментов с одной парой ног, а за ними один или несколько диплосегментов без ног; при последующих линьках на этих диплосегментах добавляются недостающие пары ног. У некоторых видов обнаружены стадии, временно имеющие по одной паре ног на 10–19 диплосегментах брюшка (Enghoff 1993). У Glyphiulus granulatus (Gervais 1847 [Julus]) и Trachyiulus tjampeanus (Attems 1903 [Cambalopsis]) обнаружена гаплоподная стадия развития (предположительно III возраст), когда позади двух первых диплосегментов брюшка, несущих по две пары ног, следуют 10–19 диплосегментов, несущих лишь по одной паре ног; в следующем возрасте на этих диплосегментах добавляется по второй паре ног (Enghoff 1993).
Как и у прочих Myriapoda, строение грудных и брюшных сегментов во многом сходно и сохраняет хорошо выраженную эволюционную корреляцию; так же сходны между собой и эволюционно коррелированы ноги всех пар, несмотря на их разное расположение. На основании того, что по строению и мускулатуре грудные сегменты сходны с брюшными диплосегментами, некоторые исследователи приходят к выводу, что каждый из этих грудных сегментов по своему происхождению также является диплосегментом, утратившем вторую пару ног; при этом половые придатки третьего туловищного сегмента считают видоизмененной второй парой ног этого диплосегмента.
(4) Строение тельсона. Тельсон [т.е. отдел тела, лежащий позади зоны нарастания новых сегментов – см. (3) и Metameria (1)] состоит из прианального кольца, обычно вытянутого назад на дорсальной стороне, пары анальных вальв, закрывающих анальное отверстие, и вентральной чешуйки (рис. 5.3C). Иногда анальная чешуйка слита с прианальным кольцом или редуцируется. У Proterandria-Nematophora от заднего края прианального кольца назад торчат 1–3 пары маленьких отчлененных грифельков (стилусов – styli) с отверстиями паутинных желез на вершине. Некоторые авторы сравнивают эти стилусы с прядильными придатками (т.н. «церками») симфил и с псевдокаудалиями (т.н. «церками») двухвосток, также имеющих протоки желез [ср.: Symphyla (8) и Prodicellura (3)]; однако в отличие от прядильных придатков симфил, стилусы диплопод сидят не на добавляющихся сегментах, а на тельсоне.
(5) Строение ног. Число члеников ноги большее, чем в других группах Atelocerata. В типичном случае нога состоит из 7 члеников, не считая претарсус [см. (7)] (рис. 5.3F); разные авторы по-разному гомологизируют эти членики с члениками ног насекомых (рис. 4.8). Кокса (1-й членик) прикреплена на вентральной стороны туловища. Коксальная аподема [см. Progoneata (2) и Collifera (5)] может отходить не от самой коксы, а от стернита латеральнее места причленения коксы (рис. 5.3F). Трохантер (2-й членик) очень короткий, кольцевидный, иногда редуцирован до полукольца или слит с коксой. Префемур, фемур, пателла и тибиа (3–6-й членики) обычно примерно равной длины. Проксимальные сочленения, включая сочленение префемура и фемура (3-го и 4-го члеников) двухмыщелковые, сочленение фемура и пателлы (4-го и 5-м члеников) и последующие одномыщелковые (Manton 1955, 1958, 1961). Тарсус (7-й членик) либо не расчленен, либо расчленен на два членика.
Плезиоморфии и изменчивые признаки Diplopoda
(6) Глаза. Обычно развиты сложные глаза; при этом они не фасеточные, а агрегированные, т.е. состоят из обособленных омматидиев (простых глаз) (рис. 5.3A, D) [см. Myriapoda (2)]. У личинки число омматидиев меньше, чем у взрослых и увеличивается с каждой линькой. У некоторых диплопод глаза полностью утрачены.
(7) Строение претарсуса. Претарсус [см. Atelocerata (3)] образует непарный коготок, часто с вентральным придатком, но без боковых коготков (в отличие от Pauropoda и некоторых других – см. Указатель признаков [2.58]).
(8) Втяжные пузырьки. Могут быть развиты втяжные пузырьки [см. Progoneata (5)]: они имеются на тазиках ног [см. (5)] у многих Nematophora и у всех Colobognatha (рис. 5.3F–G). Эти пузырьки втягиваются при помощи мышц, идущих от коксальных аподем, как и стернальные втяжные пузырьки симфил [ср.: Symphyla (6)]. У некоторых других Diplopoda имеются «коксальные органы», считающиеся гомологами втяжных пузырьков. У вымерших карбоновых Archipolypoda (см. Myriapoda: «Классификации») парные втяжные пузырьки расположены не на коксах, а между коксами, на каждом стерните (при этом имеется по два стернита на каждом диплосегменте) (Hannibal 1998).
Геологический возраст. Палеозой – ныне (рис. 2.2). К диплоподам относят некоторые ископаемые остатки, имеющие силурийский возраст. В частности, из формации Cowie в Шотландии, датируемой средним силуром, описан Cowiedesmus eroticopodus Wilson & Anderson 2004, отнесенный к Diplopoda-Proterandria, а также виды диплопод неясного систематического положения. Среди них небольшой фрагмент плохой сохранности, описанный под названием Pneumodesmus newmani Wilson & Anderson 2004; для него указано наличие дыхалец, что считается самым ранним свидетельством о появлении воздушного дыхания у животных и подтверждением гипотезы о существовании наземных биоценозов в Силуре. Однако у современных диплопод трахеи открываются не наружу, а в коксальные аподемы [см. Collifera (5)]; при этом аподемы и их внешние отверстия выполняют механическую функцию [см. Progoneata (2)], поэтому могут существовать независимо от наличия трахей и независимо от среды обитания. Несколько видов диплопод описаны также из раннего девона (Wilson & Anderson 2004). Из карбоновых и более поздних отложений известно много разнообразных диплопод.
На основании представления о том, что в силуре не было наземных биоценозов, но существовали диплоподы, некоторые авторы делают вывод о том, что наиболее древние представители таксона Diplopoda были не наземными, а первичноморскими. По другой версии таксон Diplopoda является первичноназемным, а его силурийские представители обитали в самых ранних наземных биоценозах, существовавших в силуре (см. главу II.2).
Таким образом, ошибочным является либо представление об отсутствии наземной фауны в силуре (см. главу II.2), либо представление об исходно наземном образе жизни Collifera вообще и Diplopoda-Anguiformia в частности [см. Collifera (5)], либо интерпретация ископаемых остатков C. eroticopodus как представителя Diplopoda-Anguiformia.
Состав и распространение. Около 8000 видов (Zhang 2011); всесветно.
Классификация Diplopoda. Diplopoda делятся на два голофилетических таксона – Penicillata и Anguiformia.
1. Penicillata Latreille 1825 {synn.circ.: Pselaphognatha Latzel 1884 = Podochila Bollman 1893 = Schizocephala Verhoeff 1926; nom.hier.: Polyxenus/fg1 [f:1838; g:1802]}. Тело недлинное, у разных представителей брюшко состоит из 4, 5 или 7 диплосегментов, так что всего имеется 11, 13 или 17 пар ног [см. Diplopoda (3)]; число сегментов и пар ног постоянное у каждого вида (в отличие от многих Proterandria) (Dohle 1988). Покровы мягкие, не кальцинированные (в отличие от Anguiformia). Ноги всех пар нормального строения, у самца не специализированы в качестве вторичных копулятивных органов; оплодотворение внешнее сперматофорное (в отличие от Opisthandria и Proterandria). На голове, на боках диплосегментов и на тельсоне имеются характерные пучки длинных крепких зазубренных щетинок (лишь частично показанные на рис. 5.3D). Мелкие, длина тела 2–3 мм. Около 100 видов; всесветно. Наиболее древние представители известны из нижнемелового ливанского янтаря.
2. Anguiformia Latreille 1825 {по циркумскрипции также соответствует: «Chilognatha» sensu Pocock 1887 (non Latreille 1802, nec Latzel 1884); nom.hier.: Julus/f5=g4 [f:1814; g:1758] (sine Polyxenus; incl. Glomeris)}. Кутикула кальцинирована (то же у некоторых Eucrustacea и некоторых Chelicerata). Вероятно, исходным является наличие парных ядовитых желез: у многих представителей Proterandria имеются парные ядовитые железы на всех или некоторых диплосегментах брюшка [см. Diplopoda (3)] (рис. 5.3A); у некоторых Opisthandria имеются подобные непарные дорсальные железы, которые в эмбриогенезе закладываются как парные. Anguiformia делятся на Opisthandria и Proterandria.
2.1. Opisthandria Verhoeff 1894 {synn.circ.: Onisciformes Latreille 1831 = Pentazonia Brandt 1833 = Oniscomorpha Pocock 1887; nom.hier.: Glomeris/fg1 [f:1815; g:1802] (incl. Glomeridesmus, Sphaerotherium)}. Самец имеет на одну или две пары ног больше, чем самка, и его ноги последних пар преобразованы в половые придатки – телоподы (не гомологичные гоноподам Proterandria). Оплодотворение прямое: самец, изогнувшись, забирает телоподами сперму из своих половых отверстий [находящихся позади ног II пары – см. Diplopoda (3)], а затем вводит ее в вульвы самки.
Тело недлинное, брюшко состоит из небольшого фиксированного числа сегментов. В разных таксонах бывает 7–17 диплосегментов, так что всего имеется 17–37 пар ног: у Glomeris/fg (sine Glomeridesmus, Sphaerotherium) – 17 пар у самки и 19 у самца; у Sphaerotherium/fg – 21 пара у самки и 23 у самца; у Glomeridesmus/fg – 36 пар у самки и 37 у самца (Dohle 1988). Число сегментов и пар ног постоянное у каждого вида (в отличие от многих Proterandria). Каждый диплосегмент имеет 5 обособленных склеритов: один диплотергит, пару диплоплейритов и два следующих друг за другом стернита, каждый с парой ног (отсюда название «Pentazonia»); то же у Proterandria-Colobognatha, в отличие от других Proterandria, у которых склериты каждого диплосегмента в той или иной мере слиты друг с другом. Длина тела 2–100 мм. Более 400 видов; всесветно. Известны начиная с карбона (Шаров 1962).
2.2. Proterandria Verhoeff 1894 {synn.circ.: Anguiformes Latreille 1831 (non Anguiformia Latreille 1825) = Helminthomorpha Pocock 1887 = Lophotracheata Verhoeff 1926; nom.circ.: Julus/f6=g5 [f:1814; g:1758] (sine Glomeris; incl. Polyzonium)}. У самца ноги одной или нескольких пар в передней части туловища (это ноги VIII, IX или X пары, иногда также соседние с ними) преобразованы в половые придатки, называемые гоноподы (gonopodia) (не гомологичные телоподам Opisthandria). Оплодотворение прямое: самец, изогнувшись, забирает гоноподами сперму из своих половых отверстий [находящихся позади ног II пары – см. Diplopoda (3)], а затем вводит ее в вульвы самки. У неполовозрелых самцов гоноподы или их зачатки отсутствуют.
Число диплосегментов брюшка колеблется в широких пределах и достигает 160 (в отличие от Opisthandria и Penicillata, у которых оно не бывает больше 17). У некоторых представителей число сегментов и пар ног у половозрелого животного постоянное (как и у Opisthandria, Penicillata и многих других Myriapoda, что является плезиоморфией); у многих Proterandria нарастание новых диплосегментов и ног продолжается и в половозрелом состоянии (эуанаморфоз). У некоторых Proterandria (Colobognatha и Merocheta), наоборот, на определенной стадии прекращается не только нарастание новых диплосегментов, но прекращаются и линьки (телоанаморфоз). Длина тела 2–300 мм. Около 10 000 видов (т.е. подавляющее большинство видов многоножек); всесветно. Известны начиная с карбона или ранее (см. выше).
2.2.1. Многие авторы рассматривают в составе Proterandria таксон Colobognatha Brandt 1834 {syn.circ.: Sugentia Brandt 1841; nom.typ.: Polyzonium/fg [f:1844; g:1834]}. Этот таксон характеризуется специализированным сосущим ротовым аппаратом, в котором мандибулы вестигиальные, апикальные придатки хилярия утрачены, а латеральные края хилярия более или менее слиты с головной капсулой, так что суженная передняя часть головы вместе с хилярием образуют сосущий хоботок. При этом многие Colobognatha сохраняют такую плезиоморфию, как расчленение диплосегмента на пять склеритов (рис. 5.3G) (как у Opisthandria).
2.2.2. Прочих Proterandria иногда объединяют в таксон Eugnatha Attems 1898 {nom.hier.: Julus/f7=g6 [f:1814; g:1758]} или рассматривают как несколько таксонов того же ранга, что и Colobognatha. Среди них можно упомянуть таксон Nematophora Verhoeff 1913 (non Nematophora Huxley 1851 [manuscript]) {nom.typ.: Chordeuma/f=Callipus/g [f:1847; g:1826]}, который характеризуется наличием прядильных стилусов на тельсоне [см. Diplopoda (4)].
* * *