КЛАДОЭНДЕЗИС НАСЕКОМЫХ

![]()
|
КЛАДОЭНДЕЗИС НАСЕКОМЫХ |
|||||||
|
|
|
fg | |||||
X-1.1.1. Плезиоморфон Eleuterata Fabricius 1775, или «Coleoptera» auct. – жесткокрылые, или жуки
(Somobialia![]() Eucaryota
Eucaryota![]() Metazoa
Metazoa![]() Monostomata
Monostomata![]() Haemataria
Haemataria![]() Peritoneata
Peritoneata![]() Metameria
Metameria![]()
Gnathopoda![]() Euarthropoda
Euarthropoda![]() Mandibulata
Mandibulata![]() Atelocerata
Atelocerata![]() Hexapoda
Hexapoda![]() Amyocerata
Amyocerata![]()
Pterygota![]() Metapterygota
Metapterygota![]() Neoptera
Neoptera![]() Eumetabola
Eumetabola![]() Metabola
Metabola![]() Elytrophora
Elytrophora![]() Eleuterata)
Eleuterata)
(рис. 10.2.1–10.2.7)
(рис. 10.2.1,
10.2.2, 10.2.3,
10.2.4, 10.2.5,
10.2.6, 10.2.7)
|
Названия, соответствующие по циркумскрипции: — Eleuterata Fabricius 1775; — Elytroptera Clairville 1798; — Coleopteria Rafinesque 1815; — Heteroptera Burmeister 1835 (non Heteroptera Latreille 1810); — Coleopterida Pearce 1936; — Elytraria Machatschke 1962. По циркумскрипции также соответствует: — «Coleoptera» sensu De Geer 1774 (non Linnaeus 1758). — таксонам под типифицированными названиями, образованными от родового названия Scarabaeus: Scaraboides, Scarabaeida. Иерархическое название: Scarabaeus/f3=g11 [f:1781; g:L.1758(gen.170)] (sine Xenos; incl. Cupes). |
|
Ранг. Обычно отряд, иногда надотряд.
Название. Общепринятым названием этого громадного таксона является «Coleoptera»; с начала XIX века и до настоящего времени название «Coleoptera» употребляется только применительно к таксону данной циркумскрипции. Однако это название старинное, известное из работ Аристотеля, и в прошлом оно применялось к различным таксонам, объединявшим жуков с другими насекомыми, имеющими плотные надкрылья. В 10-м издании книги Линнея «Systema Naturae», принятом за исходную точку зоологической номенклатуры, отряд Coleoptera, помимо настоящих жуков, включал роды Forficula, Blatta и Gryllus – т.е. уховерток (Dermatoptera), тараканов (Neoblattariae), богомолов (Raptoriae), палочников (Spectra) и прыгающих прямокрылых (Saltatoria) (см. I.5.4.2). Таким образом, название Coleoptera Linnaeus 1758 по своей исходной циркумскрипции не соответствует рассматриваемому здесь таксону.
Аутапоморфии Eleuterata. Особенности строения жуков рассмотрены выше, в характеристике таксона Elytrophora, объединяющего жуков (Eleuterata) и веерокрылых (Strepsiptera). Существует мнение, что Eleuterata – голофилетический таксон, отличающийся от сестринского таксона Strepsiptera некоторыми апоморфиями. В качестве таких аутапоморфий названы следующие: наличие гулы (при этом считается, что у Strepsiptera гулы не); передние крылья без видимого жилкования, в покое сомкнуты задними краями (тогда как у Strepsiptera передние крылья могут иметь жилки и в покое не смыкаются); последние сегменты брюшка втянуты внутрь (тогда как у Strepsiptera они не втянуты). В действительности ни один из этих признаков не позволяет разделить Eleuterata и Strepsiptera.
Гула имеется у всех жуков [см. Elytrophora (3)]. Утверждение об отсутствии гулы у веерокрылых высказал Кинзельбах (Kinzelbach 1971). По его мнению, имеющийся у Strepsiptera склеротизованный мост, соединяющий края головной капсулы с вентральной стороны, представляет собой не гулу, а нижнюю губу, поскольку задние тенториальные ямки находятся позади него (рис. 10.3.1A). У веерокрылых ротовой аппарат нефункционирующий, нижняя губа отсутствует, голова поперечная и сильно сокращена в длину, тенториум утрачен; вогнутости, интерпретируемые как вестигии задних тенториальных ямок, показаны лишь для вида, у которого гула имеет выемчатый задний край. Среди Eleuterata сходную форму гулы в виде моста с выемкой между задними тенториальными ямками можно встретить у представителей с вестигиальным ротовым аппаратом.
На передних крыльях жуков жилки бывают сомкнутыми и несомкнутыми, и это преобразование эволюционно обратимо [см. Elytrophora (6)]. Так что веерокрылые с несомкнутыми жилкам вполне могли произойти от жуков с сомкнутыми или несомкнутыми жилками.
Исходно у жуков последние сегменты брюшка вместе с гениталиями в покое втянуты в предыдущие сегменты [см. Elytrophora (10)]; однако у некоторых жуков, как и у веерокрылых, все сегменты брюшка утратили способность втягиваться (рис. 10.2.3A).
В настоящее время нет аргументов против предположения о том, что веерокрылые произошли от какого-то таксона в составе жуков, так что отряд Eleuterata следует считать плезиоморфоном, т.е. предположительно парафилетическим таксоном (см. Strepsiptera: «Систематическое положение»).
Размеры. Длина тела 0,2–150 мм.
Геологический возраст. Ранняя пермь – ныне (рис. 2.2). Наиболее древними представителями являются жуки, которых относят либо к современному плезиоморфону Archostemata (см. «Классификация»), либо к особым вымершим плезиоморфонам Protocoleoptera Crowson 1981 (non Protocoleoptera Tillyard 1924b) и Archecoleoptera Crowson 1981. Среди них наиболее древними являются раннепермские Tshekardocoleus/fg [fg:1944] (Пономаренко 1969, Пономаренко & Жерихин 1980, Kirejtshuk & al. 2014). Большинство видов этого таксона описаны по отдельным надкрыльям, некоторые виды известны по целым экземплярам. Раннепермские жуки имеют облик, типичный для Eleuterata: тело дорсо-вентрально уплощено, переднегрудь крупная и подвижная, передние крылья превращены в надкрылья, сходящиеся медиально по шву, заднегрудь увеличена, на брюшке лишь 6 (II–VII) видимых стернитов [см. Elytrophora (10)]. Пермские жуки отличаются от современных следующими плезиоморфиями: (1) антенны имеют 13 члеников [у современных жуков обычно 11 – см. Elytrophora (2)]; (2) на надкрыльях жилки не вполне параллельны друг другу и сохраняют некоторые развилки [см. Elytrophora (6)]. Эти признаки не позволяют провести четкую границу между пермскими и более поздними жуками, поскольку число члеников антенн известно лишь для немногих ископаемых видов, а преобразование жилок надкрыльев, возможно, происходило независимо в разных группах жуков (Пономаренко 1969).
Состав и распространение. Около 400 000 видов (Zhang 2011); всесветно.
Статус Eleuterata. Таксон Eleuterata является либо голофилетическим, либо парафилетическим, предковым для Strepsiptera (см. «Аутапоморфии»).
Классификация Eleuterata. Eleuterata делят на таксоны Adephaga, Polyphaga и Archostemata, обычно считающиеся подотрядами. Из них Adephaga и Polyphaga являются голофилетическими, тогда как Archostemata представляет собой плезиоморфон.
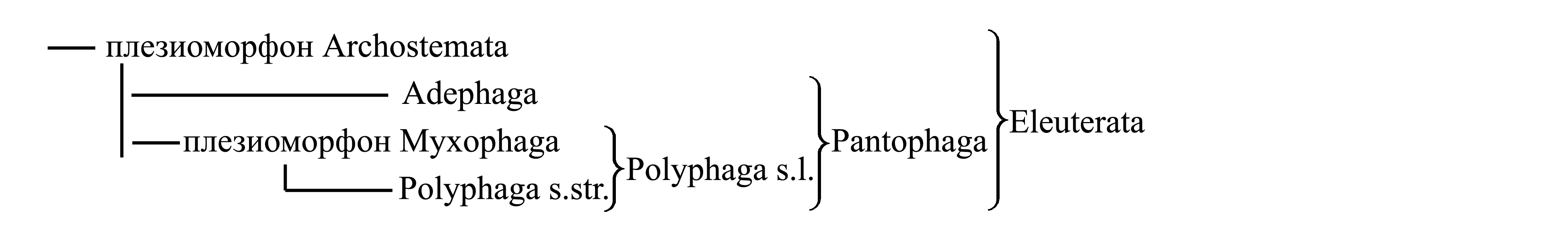
1. Archostemata Kolbe 1908 {syn.circ.: Protadephaga Kolbe 1903; nom.hier.: Cupes/fg1 [f:1857; g:1801] (incl. Omma)}. Характеризуется следующими плезиоморфиями. В отличие от Adephaga (см. ниже) у имаго задние тазики сохраняют подвижное сочленение заднегрудью. В отличие от Polyphaga (см. ниже) у имаго плейрит переднегруди не слит с трохантином и образует часть стенки тела; на заднем крыле имеется продолговатая ячейка [см. Elytrophora (7a)]; у личинки могут быть развиты ноги, имеющие отчлененную лапку и два коготка. Личинки развиваются в древесине. В современной фауне Archostemata распространены во всех частях света, но включают лишь немногим более 30 видов. Современных Archostemata принято делить на Cupes/fg2 (incl. Priacma) (около 30 видов в разных частях света) и Omma/fg1 (incl. Tetraphalerus) (несколько видов в Австралии и Южной Америке). Кроме того, к Archostemata относят много ископаемых жуков.
Дискуссии вызвало систематическое положение таксона Micromalthus/fg [f:1913; g:1878], включающего единственный вид Micromalthus debilis Leconte 1878, распространенный в Северной Америке и завезенный в южную Африку. Это мелкие мягкотелые жуки с уменьшенными надкрыльями; жилкование крыльев упрощено (в связи с чем продолговатая ячейка отсутствует). Все склериты переднегруди (тергит, плейриты и стернит) слиты без швов между ними, и криптоплейрит отсутствует; в связи с этим некоторые авторы относили Micromalthus debilis к Polyphaga на основании формального признака – отсутствия нотоплейрального шва (см. ниже). Однако у Micromalthus debilis плейрит не имеет строения, характерного для Polyphaga. Micromalthus debilis развивается в древесине и имеет очень сложный жизненный цикл, в котором сочетаются гиперметаморфоз (чередование резко различающихся по строению личиночных стадий) и педогенез (размножение личинок) (Barber 1913); помимо питающейся безногой личинки имеется расселительная личинка с ногами; у этой личинки сохраняется отчлененная лапка и два коготка (в отличие от Polyphaga s.l.).
Предположительно к Archostemata отнесен таксон Sikhotealinia/fg [fg:1996] с единственным видом Sikhotealinia zhiltzovae Lafer 1996, известным по одному экземпляру с Дальнего Востока России. Он отличается от всех прочих жуков наличием всех трех простых глазков.
Некоторые авторы относят к Archostemata таксон Crowsoniella/fg [f:1983; g:1976] с единственным видом Crowsoniella relicta Pace 1976, известным по нескольким экземплярам из Италии.
Некоторые авторы понимают таксон Archostemata широко и относят к нему наиболее древних ископаемых пермских жуков (см. «Геологический возраст»).
Вероятно, Archostemata представляют собой парафилетическую группу, предковую для других жуков. Было предложено объединять всех остальных жуков, кроме Archostemata, в таксон Pantophaga Klausnitzer 1975 {по циркумскрипции соответствует «Coleoptera» sensu Machatschke 1962}. Высказывалось ошибочное утверждение, что Pantophaga отличаются от Archostemata приобретением сплошной склеротизации надкрыльев, при которой жилки заполняют все надкрылье, так что ячейки сокращены до точек или совсем исчезли. В действительности этот признак эволюционирует обратимо и поэтому не может служить доказательством голофилии таксона [см. Elytrophora (6)]. По другой версии, Adephaga и Polyphaga произошли от разных вымерших Archostemata (Пономаренко 1969).
2. Adephaga Stephens 1828 {synn.circ.: Entomophaga Latreille 1810 (non Adephagi Clairville 1798); nom.hier.: Carabus/f1=Cicindela/g1 [f:1802; g:L.1758(gen.183)] (incl. Dytiscus)}. Характеризуется следующими аутапоморфиями.
(1) У имаго тазики задних ног [широкие, прикрепленные к заднему краю заднегруди – см. Elytrophora (5)] и первые стерниты брюшка модифицированы (рис. 10.2.2A): тазики задних ног неподвижны относительно заднегруди, направлены назад и вдавлены в стерниты брюшка таким образом, что видимая снаружи склеротизованная часть первого видимого стернита брюшка [т.е. истинного второго стернита брюшка – см. Elytrophora (8)] оказывается полностью разделенной на две доли, лежащие по бокам от тазиков (у Geodephaga-Rhysodes/fg имеется также третья доля, лежащая между тазиками); первые три видимых стернита брюшка [т.е. истинные II–IV стерниты] синсклеротичны. Уникальная апоморфия. Среди Adephaga есть как наземные жуки (Geodephaga) с бегательными ногами, так и водные жуки (Hydrocanthari), у которых задние ноги специализированы как плавательные. Высказано предположение, что общий предок Adephaga был водным, имел плавательную специализацию задних ног, и в связи с этим приобрел особое строение задних тазиков; неподвижное соединение задних тазиков с туловищем уменьшает степень свободы задних ног, но увеличивает силу гребных движений. Согласно этому предположению, у наземных представителей Adephaga задние ноги вторично утратили плавательную специализацию и снова стали бегательными.
(2) У имаго и у личинки галеа [исходно одночлениковая – см. Hexapoda (2a)] состоит из 2 пассивно-подвижных члеников. В остальном строение максиллы у имаго и у личинки сильно различается.
(3) У личинок Adephaga верхняя губа полностью слита с наличником, мандибулы утратили молу. Возможно, это связано с исходной хищной специализацией. Многие виды жужелиц перешли к фитофагии, но сохранили эти особенности строения ротового аппарата.
У Adephaga имеются следующие плезиоморфии. В отличие от Polyphaga, яичники мероистического политрофического типа [см. Eumetabola (6)]. У имаго на переднегруди сохраняются открытые плейриты (рис. 10.2.2C) (в отличие от Polyphaga). У личинок большинства Adephaga ноги развиты, с полным набором члеников [см. Hexapoda (5)]; в отличие от Polyphaga s.l., лапка [одночлениковая – см. Metabola (1Aa)] обычно отчленена от голени (лишь у некоторых сильно специализированных личинок Adephaga ноги вестигиальные, и в этом случае лапка слита с голенью); бывают развиты оба коготка [см. Pterygota (5)] (но иногда один из них уменьшен или отсутствует).
Возможно, общим для Adephaga является то, что при превращении личинки в куколку в ногах не происходит отделения гиподермы от личиночной кутикулы (в отличие от других насекомых), а в дистальных члеников личиночной ноги все ткани растворяются и не принимают участия в образовании куколочно-имагинальной ноги (в отличие от Poplyphaga) [см. Metabola (1Ccδ-2) и рис. 10.2.8].
К Adephaga относится более 40 000 видов, т.е. около 10% всех видов жуков.
Некоторые авторы выделяют в составе Adephaga следующие таксоны.
2.1. Hydrocanthari Latreille 1802b {syn.circ.: Hydradephaga MacLeay 1825; nom.hier.: Dytiscus/fg1 [f:1815; g:1758(gen.185)] (incl. Gyrinus)}. Объединяет всех водных представителей Adephaga; по мнению некоторых авторов является парафилетическим таксоном, предковым для наземных Geodephaga [см. Adephaga (1)].
2.1.1. Ephydrodephaga Kolbe 1880 {nom.hier.: Gyrinus/fg1 [f:1810; g:1762]} – вертячки. Жуки имеют особое строение ног, глаз и антенн и приспособлены к быстрому плаванию на поверхности воды. Личинки подводные, по строению брюшка напоминают Eumegaloptera: с длинными придыхальцевыми трахейными жабрами и двумя парами крючьев на пигоподе.
2.1.2. Enhydrodephaga Kolbe 1880 {nom.hier.: Dytiscus/fg2 (sine Gyrinus; incl. Haliplus). Объединяет остальных водных Adephaga; по мнению некоторых авторов является парафилетическим таксоном, предковым для наземных Geodephaga.
2.2. Geodephaga MacLeay 1825 {nom.hier: Carabus/f2=Cicindela/g2 [f:1802; g:L.1758(gen.183)] (incl. Rhysodes, Trachypachus)}. Объединяет всех наземных представителей Adephaga.
3. Polyphaga Emery 1886, s.l. {nom.hier.: Scarabaeus/f4=g12 (sine Cicindela, Cupes; incl. Sphaerius)}. Характеризуется следующими аутапоморфиями.
(1) У имаго на переднегруди трохантин слит с плейритом [не имеющим плейрального шва и несущем криптоплейрит – см. Elytrophora (4)] (рис. 10.2.4B) (в отличие от Adephaga и Archostemata, у которых трохантин переднегруди отчленен от плейрита) (Hlavac 1972, 1975).
(2) У личинки лапка всегда полностью слита с голенью, в том числе у представителей с хорошо развитыми ногами (рис. 10.2.6B–C, 10.2.7B) (в отличие от Adephaga и Archostemata, у которых одночлениковая лапка обычно отчленена от голени). Неуникальная апоморфия (см. Указатель признаков [2.49]). У некоторых Polyphaga ноги личинки вестигиальные или утрачены.
Возможно, общим для Polyphaga является то, что при превращении личинки в куколку из личиночного бедра развивается куколочно-имагинальная голень, а из личиночного тибиотарсуса – куколочно-имагинальная лапка [см. Metabola (1Ccδ-1), рис. 10.1.2B 10.2.6C–F и 10.2.7].
(3) У личинки претарсус, если развит, представляет собой один коготок (рис. 10.2.6B) (в отличие от многих Adephaga и Archostemata, у которых претарсус несет два коготка). Неуникальная апоморфия (см. Указатель признаков [2.58]).
Polyphaga s.l. делятся на Myxophaga и Polyphaga s.str.
3.1. Myxophaga Crowson 1955 {nom.typ.: Sphaerius/fg [f:1848, g:1839] (incl. Lepicerus, Hydroscapha)}. Малочисленная группа жуков, занимает промежуточное положение между плезиоморфоном Archostemata и обширным голофилетическим таксоном Polyphaga s.str. Отличается от Polyphaga s.str. плезиоморфиями: на переднегруди имаго плейрит [слитый с трохантином – см. выше, Polyphaga s.l. (1)] более или менее крупный, неподвижный, не скрыт тергитом (Hlavac 1972, 1975); заднее крыло с продолговатой ячейкой [см. Elytrophora (7)]. Myxophaga распространены во всех частях света, но включают лишь около 70 видов, ведущих водный и околоводный образ жизни.
3.2. Polyphaga Emery 1886, s.str. {nom.hier.: Scarabaeus/f5=g13 (sine Sphaerius)}. Характеризуется следующими аутапоморфиями.
(1) У имаго на переднегруди плейрит [слитый с трохантином – см. выше, Polyphaga s.l. (1)] модифицирован следующим образом (рис. 10.2.4) (Hlavac 1972, 1975). Наружная часть плейрита редуцирована, так что дорсальный мыщелок тазика находится вплотную к краю тергита (иногда скрыт нависающим краем тергита); в результате этого на переднегруди, помимо тергита и стернита [неподвижно соединенных впереди тазиков – see Elytrophora (4)] бывает видна лишь пара трохантинов, от каждого из которых внутрь тела уходит криптоплейрит; иногда трохантины также скрыты краями тергита. Такой плейрит (т.е. трохантин слитый с криптоплейритом) обычно подвижно соединен с наружной стенкой переднегруди, образованной тергитом и стернитом; у некоторых Polyphaga s.str. плейрит вторично утратил подвижность из-за того, что криптоплейрит срастается с тергитом. В отличие от Polyphaga s.str., у Myxophaga, Adephaga и Archostemata плейрит переднегруди неподвижно соединен с тергитом и стернитом, образуя часть вентро-латеральной стенки переднегруди; соответственно, у Myxophaga, Adephaga и Archostemata на вентро-латеральной стенке переднегруди бывает виден продольный терго-плейральный шов (лишь в случаях полного слияния тергита и плейрита этот шов утрачен), а у Polyphaga s.str. терго-плейрального шва не бывает.
(2) На заднем крыле ячейка oblongum отсутствует (тогда как у Myxophaga, Adephaga и Archostemata эта ячейка обычно развита, за исключением видов с вестигиальным жилкованием) [см. Elytrophora (7)].
(3) Яичники [исходно мероистические политрофические – см. Eumetabola (6)] мероистического телотрофического типа, т.е. с трофоцитами, сконцентрированными в апикальной части овариолы (в отличие от Adephaga, у которых сохраняется мероистический политрофический тип). Неуникальная апоморфия [см. Hexapoda (12)]. Этот признак исходно послужил для выделения таксона Polyphaga (Emery 1886). К настоящему времени телотрофическое строение яичников выявлено примерно для 50 видов из разных таксонов Polyphaga s.str.; представители Myxophaga и Archostemata в этом отношении не изучены, так что систематическое значение этого признака не вполне ясно (Büning 1979, 1994).
К Polyphaga s.str. относится около 90% всех видов жуков.
* * *