КЛАДОЭНДЕЗИС НАСЕКОМЫХ

![]()
|
КЛАДОЭНДЕЗИС НАСЕКОМЫХ |
|||||||
|
|
|
fg | |||||
Глава VIII. СИСТЕМАТИКА NEOPTERA: ОБЩАЯ КЛАССИФИКАЦИЯ И ОРТОПТЕРОИДНЫЕ
VIII-1. Neoptera Martynov 1923 – новокрылые
(Somobialia![]() Eucaryota
Eucaryota![]() Metazoa
Metazoa![]() Monostomata
Monostomata![]() Haemataria
Haemataria![]() Peritoneata
Peritoneata![]() Metameria
Metameria![]()
Gnathopoda![]() Euarthropoda
Euarthropoda![]() Mandibulata
Mandibulata![]() Atelocerata
Atelocerata![]() Hexapoda
Hexapoda![]() Amyocerata
Amyocerata![]()
Pterygota![]() Metapterygota
Metapterygota![]() Neoptera)
Neoptera)
(рис. 8.1–10.16.2)
|
Названия, однозначно соответствующие по циркумскрипции: — Neoptera Martynov 1923 (non Neoptera Woodworth 1906); — Neopterygota Crampton 1924; — Neoptilota Lameere 1933 (исходно Neoptilotes, во французском написании); — Opisthoptera Lemche 1940; — Neopterata Boudreaux 1979b. Иерархическое название: Pediculus/f7=Scarabaeus/fg7 [f:1777; g:L.1758(gen.170)] (sine Libellula; incl. Embia, Forficula). |
|
Ранг. Обычно от инфракласса (или отдела) до подкласса.
Название. В настоящее время общепринятым названием этого таксона является Neoptera Martynov 1923, несмотря на то, что это название преоккупировано. Ранее такое же название Neoptera Woodwort 1906 предлагалось для полифилетической группы насекомых, включающей лишь Diptera, Hymenoptera и Lepidoptera; название Neoptera Woodwort 1906 является младшим синонимом Zygothoraca Schoch 1884.
Аутапоморфии Neoptera
(1) Складывание крыльев. Имеется механизм складывания крыльев в покое, при котором крылья отводятся назад так, что их дорсальные поверхности оказываются обращенными дорсально или латерально [см. (1c)]. Это складывание крыльев обеспечивается благодаря особому строению аксиллярного аппарата [см. (1a)] и характерным чертам жилкования [см. (1b]. В связи со способностью к складыванию крыльев в ходе эволюции Neoptera неоднократно возникали сходные специализации крыльев [см. (1d)] и сходные формы редукции крыльев [см. (1e)]. В редких случаях способность складывать крылья вторично утрачена (например, у дневных бабочек – Rhopalocera).
Некоторые авторы считают способность к складыванию крыльев не апоморфией, а плезиоморфией, исходно присущей общему предку всех Pterygota; на основании этого высказывают сомнения в голофилии Neoptera и не признают этот таксон [см. (1f)] (см. Pterygota: «Классификации» IV).
(1 a) Механизм складывания крыла. В типичном случае аксиллярный аппарат [т.е. система склеритов в сочленении дорсальной поверхности крыла и тергита – см. Pterygota (4Aa)] имеет следующее строение (рис. 8.1A). Первый аксиллярный склерит (1Ax) причленяется к переднему крыловому выступу нотума; его передний отросток, изогнутый латерально, сочленяется с базисубкостой (basisubcostale) – склеритом, от которого отходит субкостальная жилка Sc [см. (1b) и Pterygota (4Ad)]; позади базисубкосты к нему подходит базирадиус (basiradiale) – склерит, от которого отходит передняя радиальная жилка RA [см. (1b) и Pterygota (4Ad)]; задняя часть 1Ax жестко связана со вторым аксиллярным склеритом. Второй аксиллярный склерит (2Ax) образует единое целое со склеритом вентральной поверхности крыла, опирающимся на плейральный столбик [см. Pterygota (4Aa)]. Третий аксиллярный склерит (3Ax) подвижно сочленен со вторым аксиллярным склеритом и с задним крыловым выступом. Дистально 3Ax подвижно сочленяется с основанием клавуса [см. (1b)].
При отведении крыла назад 3-й аксиллярный склерит переворачивается на 180°: между его основанием и задним крыловым выступом образуется вогнутая складка, а между его вершиной и основаниями сочлененных с ней жилок крыла образуется выпуклая (т.е. направленная дорсально) складка; благодаря этому переворачиванию 3-го аксиллярного склерита задняя часть основания крыла смещается медиально и надвигается на край нотума, так что все крыло поворачивается назад (рис. 8.1B). Если крыло имеет широкое основание, то при его складывании более или менее крупный участок крыла переворачивается вместе с 3-м аксиллярным склеритом (рис. 8.1C).
Далее мы будем называть этот переворачивающийся участок крыла инверталяре (invertalare), или инверталярная область (area invertalaris). Соответственно, выпуклую складку, образующуюся при складывании крыла и отделяющую инверталярную область от остальной части крыла, будем назвать инверталярной складкой (plica invertalaris). Узкую инверталярную область, не несущую жилок или включающую не более двух небольших жилок, принято называть югум (jugum), или югальная область; в этом случае жилки этой области крыла называют югальными (Ju). Исходно термин «jugum» (ярмо) был связан с ошибочным представлением о том, что у некоторых чешуекрылых югум переднего крыла обеспечивает его сцепку с задним крылом (Comstock 1918) [см. ниже, Metabola (4)]. На расширенных задних крыльях некоторых насекомых инверталярная область включает не только югум, но и лежащий впереди него участок крыла с жилками, обозначаемыми как ваннальные или анальные [см. Pterygota (4Ad)]; в таких случаях бывает трудно разобраться, какая часть инверталяре соответствует первичному югуму, а какая часть крыла вошла в состав инверталяре вторично. Мартынов (1924) предложил обозначать югальную область крыла термином неала (neala – новое крыло), а остальную часть крыла называть палеала (palaeala – древнее крыло); согласно его определению, переворачивающаяся часть крыла в некоторых случаях представляет собой «новое крыло» (neala = jugum), а в некоторых случаях, помимо нового крыла, включает часть «древнего крыла» (palaeala). Термины «неала» и «палеала» отражают представление о том, что нескладывающиеся крылья у древнекрылых (Palaeoptera) являются плезиоморфией, а способность к складыванию крыльев у новокрылых (Neoptera) – апоморфией (Мартынов 1924). Инверталярную складку (т.е. складку, по которой происходит перегибание крыла при его отведении назад), называют либо югальной, либо анальной, либо ваннальной, в зависимости от того, между какими жилками она проходит, и в зависимости от представлений о гомологии жилок и участков крыла. В отличие от терминов «югум», «неала», «ваннус», и таких названий складок, как «югальная», «анальная» и «ваннальная», термины «инверталяре» и «инверталярная складка» не зависят от гипотез о гомологии жилок и участков крыла, а лишь указывают на фактическое положение задне-проксимальной линии перегиба, образующейся при складывании крыла.
У эмбий на крыльях обеих пар инверталяре не развита [см. ниже, Idioprothoraca (5) и рис. 8.2.1A). У Rhipineoptera на передних крыльях инверталяре не развита или представлена небольшой лопастью (югумом) без жилок; на задних крыльях (ведущих при полете) инверталяре обычно занимает значительную часть крыла, несет расходящиеся жилки и складывается веерообразно; лишь у термитов и некоторых веснянок инверталяре заднего крыла редуцирована [см. ниже, Rhipineoptera (1a); Plecoptera (4) и рис. 8.3.1; Tegminoptera (3) и рис. 8.4.2A, Neoblattariae: «Разнообразие» и рис. 8.4.3; Isoptera (5) и рис. 8.4.6; рис. 8.5.5, Spectra (3) и рис. 8.6.1; Dermatoptera (5a) и рис. 8.7.1]. У Zoraptera, Panpsocoptera, Thysanoptera и Plantisuga на крыльях обеих пар инверталяре не развита (рис. 9.1.2, 9.2.1, 9.2.5, 9.2.6, 9.5.1, 9.9.1, 9.10.1, 9.11.1, 9.12.1). У цикадовых и клопов на передних крыльях инверталяре не развита; на задних крыльях (при полете сцепленных с передними) инверталяре образует крупную лопасть с одной жилкой [см. ниже, Hemelytrata (2b), рис. 9.7.1, 9.8.2]. У некоторых насекомых с полным превращением небольшая инверталяре переднего крыла (югум) имеет вид выступа и участвует в контактном аппарате [см. ниже, Metabola (4) и рис. 10.5.1, 10.9.1, 10.15.1A, 10.16.1]; у многих жуков, вислокрылок, перепончатокрылых, ручейников и чешуекрылых инверталяре заднего крыла образует более или менее крупную лопасть [см. ниже, Elytrophora (7) и рис. 10.2.2, 10.2.3; Meganeuroptera (4); Hymenoptera (5b).] и рис. 10.7.1, 10.7.2B; Trichoptera (5) и рис. 10.15.1B; Neolepidoptera : «Классификация»: Frenatae]; у двукрылых инверталяре, если развита, то представляет собой верхний калиптер [см. ниже, Diptera (2b)]; у некоторых насекомых с полным превращением инверталяре не развита на крыльях обеих пар.
(1 b) Строение крыла. Область крыла, лежащая впереди от инверталярной складки и не переворачивающаяся в покое, разделена клавальным желобом на преклавус и клавус (рис. 8.1A).
Клавальный желоб (impressio clavalis) представляет собой вогнутость с дорсальной стороны крыла и, соответственно, выпуклость с вентральной стороны; он тянется от основания крыла к его заднему краю, бывает прямым (в частности у Arthroidignatha), либо изогнут. Точка впадения клавального желоба в задний край крыла называется задний узелок, или задний нодус (nodus posterior). Клавальный желоб либо не имеет жилок, либо по его дну проходит продольная жилка (считающаяся при различных интерпретациях либо Cu2, либо Pcu, либо новообразованием), либо вдоль него проходит одна или две жилки: спереди – Cu2, сзади – Pcu. Клавальный желоб называют также клавальной, или кубитальной складкой (поскольку он расположен позади кубитальной жилки); однако слово «складка» здесь представляется неудачным, поскольку, в отличие от инверталярной складки [см. (1a)], по клавальному желобу не производится складывания крыла; клавальный желоб постоянно вогнут и может обеспечивать лишь небольшую подвижность.
У некоторых Neoptera, помимо заднего нодуса, имеется передний нодус (nodus anterior) – перерыв костальной жилки (рис. 9.9.1A, 10.7.1), от которого поперек крыла к заднему нодусу тянется нодальная линия – борозда, обеспечивающая гибкость крыла.
Преклавус (praeclavus), или преклавальная область обычно имеет следующие жилки. Помимо идущей по переднему краю C и идущих вдоль него и обычно неветвящихся вогнутой Sc и выпуклой RA [см. Pterygota (4Ad)], имеются жилки RS, M, Cu1 и Cu2. У разных новокрылых насекомых жилки RS, M, Cu1 и Cu2 бывают выпуклыми, либо вогнутыми, либо нейтральными; бывают ветвящимися или неветвящимися; в основании они бывают различным образом соединены друг с другом и/или с RA. Иногда те или иные жилки утрачены. Около самого основания жилки C и Sc могут быть жестко связаны крепкой плечевой жилкой (vena humeralis – h); в отличие от костальной скобы поденок, плечевая жилка новокрылых является поперечной и сходна с плечевой жилкой стрекоз. Поскольку Sc и RA посредством 1-го и аксиллярного склерита сочленены с передним крыловым выступом нотума [см. (1a)], а остальные жилки преклавуса так или иначе связаны с RA, можно считать, что весь преклавус связан с передним крыловым выступом.
Иногда вместо термина «преклавус» используют термин «ремигиум» (remigium), что означает «весло» и связано с его активным движением при полете (Snodgrass 1935); однако исходно термин «ремигиум» используется лишь для проксимального участка радиальной жилки в крыле двукрылых (Lowne 1890–1892) [см. ниже, Diptera (2b)].
Клавус (clavus), или клавальная область, посредством 3-го аксиллярного склерита сочленен с задним крыловым выступом нотума [см. (1a)]. Жилки клавуса начинаются от его места сочленения с 3-м аксиллярным склеритом; они называются либо анальными (1A–3A) (Comstock & Needham 1898, Comstock 1918a), либо посткубитальной (Pсu) и ваннальными (V1 и V2) (Snodgrass 1935).
Исходно термин «клавус» использовался для соответствующей области на передних крыльях клопов (рис. 9.8.2D) (Fieber 1844). Иногда вместо термина «клавус» используют термин «ваннус» (vannus); однако исходно термин «ваннус» используется как для клавуса, так и для инверталярной области крыла (Snodgrass 1935).
(1 c) Способы складывания крыльев. У разных новокрылых насекомых различают следующие типы складывания крыльев.
Кровлеобразное (рис. 8.1F): костальный край крыла направлен вентрально или вентро-латерально, а дорсальная поверхность крыла обращена латерально или латеро-дорсально; крылья одной пары не налегают друг на друга, их плоскости лежат под углом друг к другу. Кровлеобразное складывание свойственно многим (но не всем) представителям таксонов Eupsocida, Auchenorrhyncha, Saltipedes (рис. 9.9.2C), Scytinelytra (рис. 9.10.3G), Gynaptera, Neuropteroidea, Nannomecoptera, Metamecoptera, Amphiesmenoptera и некоторых других.
Плоское (рис. 8.1D): костальный край направлен латерально, дорсальная поверхность обращена дорсально; крылья лежат в одной плоскости, крылья одной пары могут налегать или не налегать друг на друга; при налегании одно из крыльев (левое или правое) может занимать постоянное верхнее положение. Плоское складывание свойственно Embioptera, Zoraptera, Thysanoptera, Heteropteroidea (рис. 9.8.1A), Hymenoptera [см. ниже, Hymenoptera (6)], Scorpiomuscae и некоторым другим.
Облегающее (рис. 8.1E): при складывании крыло сгибается вдоль так, что его передняя часть расположена сбоку от тела как при кровлеобразном складывании, а его задняя часть лежит на дорсальной стороне тела как при плоском складывании (рис. 8.5.4B); при этом у крыльев одной пары задние части могут налегать или не налегать друг на друга. Продольное сгибание крыла происходит по-разному в разных таксонах (см. Указатель признаков [2.104–2.105]).
Применительно к надкрыльям неподвижно согнутую переднюю часть называют эпиплевра (epipleura); при сложенных крыльях эпиплевра сбоку налегает на плейрит (рис. 8.7.1B). Этот же термин «эпиплевра» можно использовать и применительно к соответствующей части личиночного протоптерона в случаях, когда притоптерон направлен назад и лежит на спине, а его костальный край загнут и сбоку налегает на плейрит (рис. 8.2.2C).
Дополнительные продольные складки. У некоторых насекомых переднее или заднее крыло приобрело способность при складывании перегибается не только по инверталярной складке, но и по дополнительным продольным складкам, становясь значительно ýже, чем в расправленном состоянии.
Дополнительные поперечные складки. В некоторых далеких друг от друга таксонах насекомых крылья приобрели способность складываться поперек; благодаря этому крыло, будучи длиннее тела, в сложенном состоянии становится короче и полностью умещается на спине насекомого. Поскольку внутри крыла ни мышц, ни сухожилий, ни аподем никогда не бывает, поперечное складывание крыла во всех случаях происходит только за счет того, что при отведении крыла назад на нем образуются особые продольные складки, которые вызывают образование определенных поперечных складок, сгибающих крыло. Среди переднемоторных насекомых способность к поперечному складыванию передних крыльев имеется у полушаровидных щитников [см. ниже, Heteropteroidea (3)]. Способность к поперечному складыванию задних крыльев независимо возникла у некоторых заднемоторных насекомых – у жуков, уховерток и некоторых тараканов [см. ниже, Neoblattariae: «Разнообразие»; Dermatoptera (5a); Elytrophora (7)].
(1 d) Пропорции и форма крыльев. Среди новокрылых имеются как бимоторные, так и переднемоторные, и заднемоторные насекомые. Вероятно, здесь, как и в целом для насекомых, бимоторность первична [см. Pterygota (4F)]. У большинства бимоторных новокрылых насекомых задние крылья, будучи сходны с передними, в то же время короче передних настолько, что при отведении назад и складывании крыльев на спине вершины передних и задних крыльев совпадают. Это совпадение проявляется только в позе покоя у имаго, тогда как у личинки вершины задних протоптеронов (т.е. зачатков задних крыльев) могут выступать из под передних. Неясно, каким образом такое совпадение очертаний крыльев возникало и поддерживалось в ходе эволюции: оно не связано с особенностями органогенеза (поскольку проявляется только на заключительной стадии онтогенеза после окончательного формирования крыла) и не имеет значения для естественного отбора (поскольку у первичнобимоторных новокрылых передние крылья не служат защитными органами).
У переднемоторных новокрылых задние крылья, как правило, еще сильнее уменьшены и в покое не достигают вершин передних.
Помимо бимоторности и переднемоторности (свойственных как древнекрылым, так и новокрылым), среди новокрылых встречается заднемоторность, при которой передние крылья, в покое закрывающие задние, служат преимущественно или исключительно для зашиты задних крыльев. Заднемоторность независимо возникла у Tegminoptera и Elytrophora. Исходно у заднемоторных насекомых соотношение длин передних и задних крыльев такое же, как у бимоторных новокрылых (при этом бóльшая площадь задних крыльев достигается только благодаря их большей ширине за счет веерообразно складывающиеся инверталярной области); у некоторых заднемоторных насекомых пропорции изменены, и задние крылья бывают значительно длиннее передних; в этих случаях они в покое либо выступают из под передних, либо приобрели особые механизмы для поперечного складывания [см. (1c)].
Среди Neoptera бимоторность с одинаковым строением переднего и заднего крыльев имеется только у эмбий [см. ниже, Idioprothoraca (5)]. У Rhipineoptera заднее крыло исходно отличается от переднего наличием анального веера, и крылья в той или иной мере дифференцированы в направлении заднемоторности [см. ниже, Rhipineoptera (1)]. У Parametabola крыловой аппарат переднемоторный [см. ниже, Parametabola (2)]. У насекомых с полным превращением передние и задние крылья исходно дифференцированы, но встречаются все типы крылового аппарата – бимоторный, переднемоторный и заднемоторный [см. ниже, Metabola (4)].
(1 e) Случаи редукции крыльев. Поскольку новокрылые насекомые способны складывать крылья так, что они не мешают им вести нелетающий образ жизни, у многих из них происходит редукция крыльев и утрата способности к полету [в то время как среди древнекрылых утрата крыльев неизвестна, а уменьшение крыльев встречается редко – см. Pterygota (4F)]. В разных случаях редукции крыльев особенности строения птероторакса, исходно свойственные крылатым насекомым [см. Pterygota (4Ab) и (4Ac)], либо утрачиваются, либо в той или иной мере сохраняются. В некоторых случаях вестигии крыльев представляют собой уменьшенные и упрощенные имагинальные крылья – то есть имеют сочленение с туловищем и занимают такое же положение, как сложенные крылья у полнокрылых форм. В других случаях вестигии крыльев у имаго сохраняют строение личиночных протоптеронов – то есть представляют собой выступающие края нотума [см. ниже, Idioprothoraca (3), Dermatoptera (5e)]. У некоторых видов имеются как полнокрылые формы, способные к полету, так и бескрылые и/или короткокрылые нелетающие формы; те и другие формы бывают обоих полов, либо одного пола. Часто встречается такой половой диморфизм, при котором у самцов крылья вполне развиты, а у самок нефункционирующие или утрачены; у некоторых насекомых, наоборот, самки могут быть полнокрылыми, а самцы нелетающими.
У эмбий крылья отсутствуют у самок, а у самцов имеются или отсутствуют [см. ниже, Embioptera (1d)]. У гриллоблаттид крылья отсутствуют [см. ниже, Notoptera (1)]. У веснянок крылья бывают развиты, укорочены или отсутствуют; иногда развиты только у части особей или только у самок [см. ниже, Plecoptera (1E)]. У различных видов в составе Raptoriae и Neoblattariae крылья бывают развиты, укорочены или отсутствуют; иногда развиты только у части особей или только у самцов. У термитов плодовитая каста крылатая, бесплодная бескрылая [см. ниже, Isoptera (1)]. У различных видов Saltatoria, Spectra и Dermatoptera крылья бывают развиты, укорочены или отсутствуют; иногда развиты только у части особей или только у самцов. У зораптер каждый вид имеет крылатую и бескрылую формы [см. ниже, Zoraptera (1)]. У сеноедов (Parapsocida и Eupsocida) крылья бывают развиты, укорочены, отсутствуют или развиты только у самцов. У всех Parasita крылья отсутствуют [см. ниже, Parasita (9)]. У Thysanoptera и Hemelytrata (Auchenorrhyncha, Coleorrhyncha и Heteroptera) крылья бывают развиты, укорочены или отсутствуют; иногда развиты только у части особей или только у самцов. У листоблошек и белокрылок (Psyllaleyroda) крылья развиты. У каждого вида тлей крылья развиты у части форм, а у части партеногенетических самок отсутствуют [см. ниже, Gynaptera (1)]. У кокцид крылья развиты у самцов и отсутствуют у самок [см. ниже, Gallinsecta (1)]. У жуков крылья бывают развиты, укорочены или отсутствуют [см. Elytrophora (6), (7)]. У веерокрылых крылья развиты у самцов и отсутствуют у самок [см. ниже, Strepsiptera (2) и (3)]. У Neuropteroidea крылья развиты. У перепончатокрылых (Hymenoptera) крылья обычно развиты, иногда укорочены, отсутствуют или развиты только у самцов; у муравьев крылья развиты у плодовитой касты и отсутствуют у бесплодной касты. Среди скорпионниц у Nannomecoptera крылья развиты; у Metamecoptera и Scorpiomuscae крылья бывают развиты, укорочены, отсутствуют или развиты только у самцов; у бореид крылья особым образом укорочены [см. ниже, Raphioptera (3)]. У блох крылья полностью утрачены [см. ниже, Aphaniptera (1a)]. У Diptera, Trichoptera и Lepidoptera крылья обычно развиты, иногда укорочены, отсутствуют или развиты только у самцов.
(1 f) Альтернативная интерпретация неоптерии. Некоторые исследователи придерживаются другой гипотезы об эволюции крыльев, согласно которой крылья изначально были способны складываться на спине, а Ephemeroptera, Odonata и некоторые вымершие группы утратили эту способность в связи со специализацией (Расницын 1976, 1980). Иными словами, эти авторы считают Neoptera парафилетической группой, предковой для Palaeoptera, и на основании этого не признают таксон Neoptera (см. Pterygota: «Классификации» VIII). В пользу гипотезы о том, что неспособность складывать крылья у Palaeoptera является вторичной, может свидетельствовать тот факт, что у всех Ephemeroptera в основании крыла задний выступ базальной пластинки (образующий подвижное сочленение с AxP) отделен от остальной части базальной пластинки бороздой [см. Ephemeroptera (7b) и рис. 7.2.4A: pbp]; существует предположение, что этот задний сочленовный выступ представляет собой третий аксиллярный склерит, неподвижно слившийся с базальной пластинкой (Бродский 1974).
Признаки Neoptera неясного филогенетического значения
(2) Форма птероторакса. Плейриты птероторакса [средне- и заднегруди – см. Pterygota (4)] и их плейральные швы [исходно дорсовентральные – см. Pterygota (3)] либо не наклонены, либо наклонены дорсальной стороной вперед, а вентральной назад, так что основания ног находятся на одном уровне или позади оснований крыльев того же сегмента (в отличие от Odonata, у которых птероторакс наклонен в обратную сторону).
(3) Утрата пателло-тибиального шва. Пателло-тибиальный шов [см. Hexapoda (5)] всегда утрачен, может сохраняться лишь вдавление на наружной стороне голени вблизи коленного сочленения. То же у Entognatha, Triplura и некоторых Ephemeroptera.
(4) Шпоры. Каждая голень обычно с парой вершинных (апикально-внутренних) шпор; каждая шпора (calcar) представляет собой отчлененный придаток, обычно склеротизованный и имеющий шиповидную форму. Внешне шпора похожа на крепкую заостренную щетинку; в отличие от настоящей щетинки [образованной только сенсорными и оболочечными клетками – см. Euarthropoda (5a)], шпора имеет внутри многоклеточную гиподерму и при линьке насекомого линяет, а не образуется заново; поэтому перед линькой растущая шпора остается вставленной в старую кутикулу шпоры хотя бы своей вершиной (как на рис. 8.4.4C); соответственно, в отличие от щетинки, шпора может нести щетинки или их производные. Обе шпоры сидят на вершине голени, на ее вентральной стороне – т.е. на внутренней стороне по отношению к коленному сочленению; у тех насекомых, у которых лапка резко отгибается наружу, шпоры оказываются торчащими и могут использоваться для лучшего сцепления ноги с субстратом; но у некоторых насекомых, имеющих шпоры, лапка неспособна отгибаться, и шпоры не выполняют этой функции.
Шпоры имеются у многих Rhipineoptera и у большинства имаго и куколок Metabola, но утрачены у всех Parametabola и в некоторых других таксонах; иногда шпоры утрачены лишь на голенях некоторых пар, иногда голень несет лишь одну шпору. Иногда, наоборот, число шпор увеличено, и они могут располагаться не только на вершине голени, но и на других местах голени и бедра (см. Указатель признаков [2.48]). Иногда на месте утраченных шпор имеются сходные по строению и функциям образования, но другой природы – зубцы, крепкие щетинки и др.
У имаго и личинок Embioptera шпоры отсутствуют (рис. 8.2.1A). У имаго и личинок Notoptera на каждой голени имеются две вершинные шпоры (рис. 8.2.3A). У имаго и личинок большинства Plecoptera на каждой голени имеются две небольшие вершинные шпоры (рис. 8.3.1A, 8.3.2A), иногда лишь одна шпора, либо шпоры утрачены (Zwick 2000). У Pandictyoptera на каждой голени имеются две вершинные шпоры; у имаго и старших личинок тараканов (Palaeoblattariae и Neoblattariae), кроме того, имеются дополнительные шпоры, разбросанные по всей голени [см. ниже, Pandictyoptera (5), рис. 8.4.4B]. У имаго и личинок Saltatoria, помимо двух вершинных шпор, на голенях прыгательных задних ног обычно бывают развиты дополнительные шпоры [см. ниже, Saltatoria (2a) и рис. 8.5.5A]. У имаго и личинок палочников и уховерток шпоры утрачены [см. ниже, Spectra (5) и Dermatoptera (6)]. У всех Parametabola вершинные шпоры голени отсутствуют, могут быть развиты лишь вторичные шпоры [см. ниже, Parametabola (4)]. У всех личинок Metabola шпоры отсутствуют [см. Metabola (1Cc)]. У имаго большинства Metabola на каждой голени имеются две вершинные шпоры; у Strepsiptera, Aphaniptera и в отдельных таксонах в составе Eleuterata, Hymenoptera, Diptera и Lepidoptera на ногах тех или иных пар или на всех ногах одна или обе вершинные шпоры утрачены. У имаго некоторых перепончатокрылых, ручейников и бабочек, помимо апикально-внутренних шпор, бывают развиты похожие на них срединно-внутренние шпоры голени [см. ниже, Hymenoptera (12) и Amphiesmenoptera (7), рис. 10.7.1 и 10.16.1].
Предположение о том, что наличие двух апикально-внутренних шпор голени является аутапоморфией Neoptera, делается на основании того, что у других насекомых (Odonata, Ephemeroptera, Triplura и Entognatha) подобные шпоры не встречаются: среди этих таксонов только у Zygentoma имеется отчлененный зубцевидный вырост на вершине голени, но он расположен не на внутренней, а на наружной стороне голени, всегда один, и не имеет щетинкоподобного строения [ср.: Zygentoma (3)].
Геологический возраст. Середина карбона – ныне. Древнейшие представители Neoptera известны из намюра и относятся к числу наиболее древних известных крылатых насекомых, наряду с Protorrhynchota и Odonatoptera (Brauckmann, Brauckmann & Groning 1996). Из намюра описаны новокрылые насекомые Ampeliptera limburgica Pruvost 1927, Stygne roemeri Handlirsch 1906, Protoprosbole straeleni Laurentiaux 1952.
Статус Neoptera. Некоторые авторы не признают таксон Neoptera, поскольку считают его парафилетическим (см. Pterygota: «Классификации» VIII), а способность складывать крылья считают плезиоморфией [см. Neoptera (1f)].
Классификации Neoptera.
I. Деление Neoptera на Polyneoptera, Paraneoptera и Oligoneoptera. Исходно (Мартынов 1923, 1938) Neoptera были разделены на три подчиненных таксона равного ранга (когорты) – Polyneoptera, Paraneoptera и Oligoneoptera. Названия этих когорт были связаны с тем, что у Polyneoptera на заднем крыле неала [см. Neoptera (1a)] имеет много жилок, у Paraneoptera неала либо имеет одну жилку, либо не развита, а у Oligoneoptera неала часто бывает без жилок или не развита.
Таксон Polyneoptera Martynov 1923 {synn.circ.: Diplomerata Börner 1904a (non Diplomerata Kirby 1892) = Orthopteradelphia Crampton 1924} включает эмбий (Embioptera), гриллоблаттид (Notoptera) и насекомых, объединяемых нами в таксон Rhipineoptera. Общими признаками Embioptera, Notoptera и Rhipineoptera являются лишь плезиоморфии: ротовой аппарат грызущий, максиллярные щупики 5-члениковые, лабиальные щупики 3-члениковые [см. Hexapoda (2с) и (3с)] (название Diplomerata связано с такой гомологизацией члеников щупика с члениками ног, при которой «femur» максиллярного щупика считался двучлениковым); имеются церки [только парные – см. Metapterygota (4)]; метаморфоз не усложнен, сохраняется значительное сходство личинок и имаго. В характеристике таксона Polyneoptera указывалось, что имеется тенденция к заднемоторности, а задние крылья имеют анальный веер или следы его редукции. На самом деле наличие анального веера можно интерпретировать как аутапоморфию Rhipineoptera [см. ниже, Rhipineoptera (1)]. У Embioptera этот признак отсутствует: в отличие от всех Rhipineoptera, их крылья не только не имеют анального веера или каких-либо его вестигиев, но и не имеют никаких следов заднемоторной специализации [см. ниже, Idioprothoraca (5)].
Таксон Paraneoptera является голофилетическим; его более однозначным названием является Parametabola (см. ниже, Parametabola).
Таксон Oligoneoptera также является голофилетическим; его старшим названием является Metabola (см. ниже, Metabola).
II. Деление Neoptera на Idioprothoraca, Rhipineoptera и Eumetabola. В связи с тем, что голофилия таксона Polyneoptera никак не обоснована, его следует рассматривать как два не связанных между собою таксона – (1) Idioprothoraca (объединяющий Embioptera и Notoptera) и (2) Rhipineoptera (объединяющий всех Polyneoptera кроме Embioptera и Notoptera). Таксоны Parametabola (= Paraneoptera) и Metabola (= Oligoneoptera) имеют синапоморфии, позволяющие объединить их в таксон Eumetabola. Таким образом, вместо трех когорт, предложенных А.В. Мартыновым (см. I), Neoptera следует делить на три других подчиненных таксона – Embioptera, Rhipineoptera и Eumetabola (Kluge 2012).
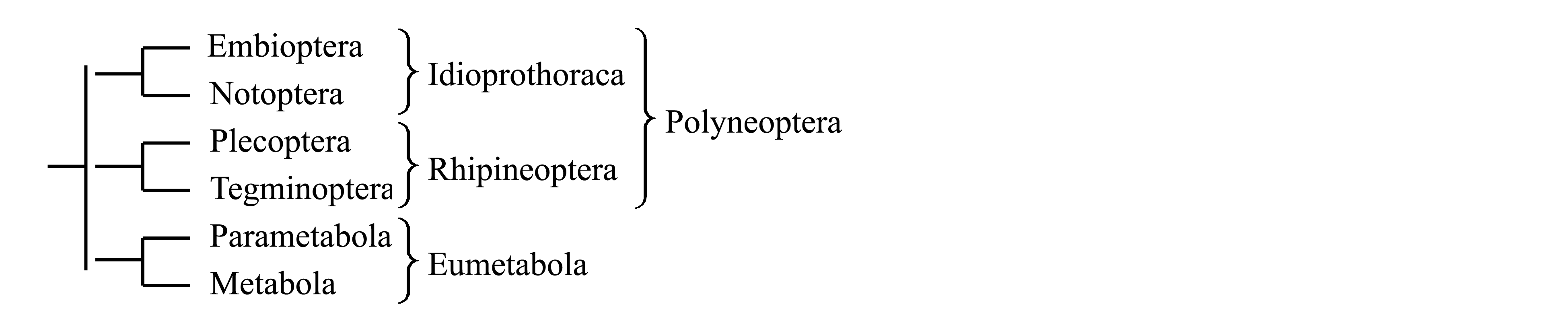
III. Другие классификации. В настоящее время предложены классификации, в которых таксон Polyneoptera расформирован, а некоторые из входивших в него групп объединены с Eumetabola. В частности Büning (1996) предложил на основе особенностей оогенеза выделить таксон Promeroista, объединяющий Plecoptera и Meroista; в свою очередь, таксон Meroista объединяет Dermatoptera с Acercaria и Metabola (см. ниже, Plecoptera: «Систематическое положение» V; Dermatoptera: «Систематическое положение» I). Некоторые авторы не признают таксон Parametabola (см. ниже, Zoraptera: «Систематическое положение» II–V).
Здесь мы принимаем деление Neoptera на 3 подчиненных таксона – Idioprothoraca, Rhipineoptera (стр. 0000) и Eumetabola (cтр. 0000).
Neoptera, или Pediculus/f7=Scarabaeus/g7
![]() 1. Idioprothoraca, или Embia/fg1
1. Idioprothoraca, или Embia/fg1
![]() 1.1. Embioptera, или Embia/fg2
1.1. Embioptera, или Embia/fg2
![]() 1.2. Notoptera, или
Grylloblatta/fg1
1.2. Notoptera, или
Grylloblatta/fg1
![]() 2. Rhipineoptera, или
Gryllus/f1=Forficula/g1
2. Rhipineoptera, или
Gryllus/f1=Forficula/g1
![]() 2.1. Plecoptera, или Perla/fg1
2.1. Plecoptera, или Perla/fg1
![]() 2.2. Tegminoptera, или
Gryllus/f2=Forficula/g2
2.2. Tegminoptera, или
Gryllus/f2=Forficula/g2
![]() 2.2.1. Pandictyoptera, или
Blatta/fg1
2.2.1. Pandictyoptera, или
Blatta/fg1
![]() 2.2.2. Saltatoria, или
Gryllus/f3=g1
2.2.2. Saltatoria, или
Gryllus/f3=g1
![]() 2.2.3. Spectra, или Phasma/fg1
2.2.3. Spectra, или Phasma/fg1
![]() 2.2.4. Dermatoptera, или
Forficula/f1=g3
2.2.4. Dermatoptera, или
Forficula/f1=g3
![]() 3. Eumetabola,
или Pediculus/f8=Scarabaeus/g8
3. Eumetabola,
или Pediculus/f8=Scarabaeus/g8
![]() 3.1. Parametabola,
или Pediculus/f9=Cicada/g1
3.1. Parametabola,
или Pediculus/f9=Cicada/g1
![]() 3.2. Metabola, или
Scarabaeus/f1=g9
3.2. Metabola, или
Scarabaeus/f1=g9