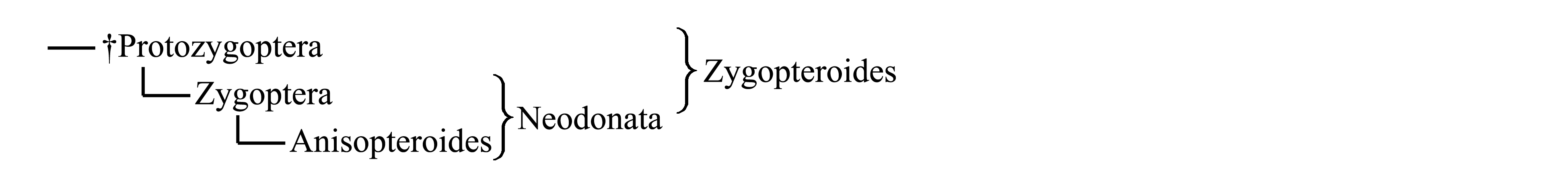КЛАДОЭНДЕЗИС НАСЕКОМЫХ

![]()
|
КЛАДОЭНДЕЗИС НАСЕКОМЫХ |
|||||||
|
|
|
fg | |||||
VII-1.2.1.B-1. Odonatoptera Lameere 1900, или
Odonata s.l.
(Somobialia![]() Eucaryota
Eucaryota![]() Metazoa
Metazoa![]() Monostomata
Monostomata![]() Haemataria
Haemataria![]() Peritoneata
Peritoneata![]() Metameria
Metameria![]()
Gnathopoda![]() Euarthropoda
Euarthropoda![]() Mandibulata
Mandibulata![]() Atelocerata
Atelocerata![]() Hexapoda
Hexapoda![]() Amyocerata
Amyocerata![]()
Pterygota![]() Metapterygota
Metapterygota![]() Odonata
Odonata![]() – Odonatoptera)
– Odonatoptera)
(рис. 7.3.1–7.3.6)
(рис. 7.3.1,
7.3.2, 7.3.3,
7.3.4, 7.3.5,
7.3.6)
|
Названия, однозначно соответствующие по циркумскрипции: — Odonatoptera Lameere 1900 (исходно Odonatopteres, во французском написании); — Tylopteradelphia = Pantyloptera Crampton 1924; — Panaeshnoptera Crampton 1928a; — Odonatoidea Lameere 1936 (исходно Odonatoides, во французском написании); — Odonatomorpha Demoulin 1970; — Odonatopterata Boudreaux 1979b; — Eupalaeoptera Carle 1982; — Odonatopteroida Müller 1978. Названия, неоднозначно соответствующие по циркумскрипции (соответствующие также Neodonata) – см. выше, Odonata. По циркумскрипции также соответствует таксонам под типифицированными названиями, образованными от родового названия Libellula – Libellulida, Libellulones и др. Иерархическое название: Libellula/fg1 [f:1781(ord.5); g:1758(gen.207)] (incl. Meganeura, Triadophlebia, Kennedya, Ditaxineura). |
|
Ранг. Обычно от отряда до инфракласса.
Аутапоморфии, достоверно относимые к Odonatoptera (о прочих аутапоморфиях см. выше, Odonata)
(1) Модификация груди [см. Odonata (4)]. Плейриты средне- и заднегруди увеличены и скошены таким образом, что эпистерны среднегруди сближены или сомкнуты на дорсальной стороне, а эпимеры заднегруди сближены или сомкнуты на вентральной стороне; в результате этого основания ног смещены вперед, а основания крыльев – назад (рис. 7.3.1E). Уникальная апоморфия.
(2) Стебельчатое крыло [см. Odonata (6)]. В проксимальной части крыло либо сужено и имеет вид стебелька, ограниченного сзади анальной жилкой (A) и имеющего уменьшенное число продольных жилок (рис. 7.3.2, 7.3.5E–F), либо не сужено, но и в этом случае в проксимальном участке, соответствующем стебельку, число продольных жилок уменьшено (рис. 7.3.5A–D, 7.3.6H); жилки RS и M отходят от RA там, где кончается стебелек; позади A нет жилок, отходящих от корня крыла.
Плезиоморфии и признаки неясного филогенетического значения, достоверно относимые к Odonatoptera (о прочих признаках см. выше, Odonata)
(3) Древнекрылость [см. Odonata (6)]. Крылья неспособны складываться на спине. То же у Panephemeroptera и Protorrhynchota. Вероятно, плезиоморфия, в отличие от Neoptera; по мнению некоторых авторов — апоморфия (см. Pterygota: «Классификации» IV).
(4) Чередование выпуклых и вогнутых жилок [см. Odonata (6)]. На крыле чередуются выпуклые и вогнутые продольные жилки: позади выпуклой C, вогнутой Sc и выпуклой RA (свойственных всем Pterygota), имеется начинающаяся общим стволом и далее разветвляющаяся вогнутая жилка RS, позади нее выпуклая M, вогнутая CuP и выпуклая A (рис. ). То же у Panephemeroptera и Protorrhynchota, что может расцениваться как симплезиоморфия или синапоморфия (см. Pterygota: «Классификации» IV).
(5) Две триады RS [см. Odonata (6)]. Жилка RS [вогнутая – см. (4)] разветвляется, образуя триаду с двумя вогнутыми ветвями RSa и RSp и выпуклой интеркалярной жилкой iRS между ними; передняя ветвь этой триады разветвляется, образуя еще одну триаду с двумя вогнутыми ветвями RSa1 и RSa2 и выпуклой интеркалярной iRSa (рис. 7.3.2A). То же у Panephemeroptera, что может расцениваться как симплезиоморфия или синапоморфия (см. Ephemeroptera: «Систематическое положение» II).
(6) Бимоторность. Крылья всегда хорошо развиты, удлиненные, передние и задние крылья примерно равной длины; в полете крылья работают независимо (не сцепляясь друг с другом), так что все Odonatoptera бимоторные, перехода к переднемоторности не происходит.
Геологический возраст. Карбон – ныне (о древнейших представителях – см. ниже, Палеозойско-триасовые стрекозы).
Классификации Odonatoptera. Подавляющее большинство современных стрекоз относится к двум крупным таксонам – Zygoptera и Anisoptera. Каждый из этих таксонов характеризуется комплексом признаков, но полярность этих признаков вызвала дискуссии. С достаточной уверенностью можно считать апоморфиями по крайней мере два признака Anisoptera: разделение дискоидальной ячейки на треугольник и надтреугольник (рис. 7.3.6G–H) и наличие у самцов выроста эпипрокта (рис. 7.3.4E). Для большинства Zygoptera характерна своеобразная форма крыла со стебельком, ограниченным сзади анальной жилкой (A), и с проксимальным положением узелка (рис. 7.3.2A). Помимо Zygoptera, такая форма крыла имеется у вымерших палеозойско-триасовых таксонов Protozygoptera и Triadophlebioptera (рис. 7.3.5E–F). В то же время некоторые из синапоморфий Zygoptera и Anisoptera [см. ниже, Neodonata (1)–(6)] у названных палеозойско-триасовых стебельчатокрылых стрекоз отсутствуют. Такое распределение признаков позволяет предположить, что наиболее примитивным в современной фауне является таксон Zygoptera, произошедший от пермско-триасовых стебельчатокрылых стрекоз – точнее, от Protozygoptera, крылья которых имеют такое большое сходство с крыльями Zygoptera, что некоторые авторы объединяют их в таксон Zygoptera s.l., или Zygopteroides. Согласно этой гипотезе таксон Anisopteroides (включающий в себя Anisoptera) произошел от Zygoptera:
В пользу того, что крылья Anisoptera, не имеющие стебелька и с узелком, удаленным от основания, произошли от стебельчатых крыльев Zygoptera, свидетельствуют следующие особенности крыльев Anisoptera: (1) в проксимальной части крыла позади A нет первичных анальных жилок, идущих от корня крыла (рис. 7.3.6H), что можно истолковать как вторичное расширение крыла позади стебелька; (2) среди множества поперечных предузелковых жилок обычно заметны две первичные предузелковые (более толстые, чем остальные) (рис. 7.3.6H: an1, an2) – отсюда можно сделать вывод, что у предков Anisoptera узелок находился значительно проксимальнее и перед ним помещались только две поперечные жилки.
Помимо приведенной гипотезы высказывались другие, в которых предполагается неоднократное возникновение некоторых признаков. К ним относятся гипотезы о независимом происхождении Zygoptera и Anisoptera (т.е. о полифилии Neodonata):

и гипотезы о происхождении Zygoptera из Anisoptera:
Эти гипотезы в значительной мере связаны с представлением о том, что Odonata произошли от Palaeodictyoptera-подобных предков (раньше предполагали даже непосредственное происхождение Odonata от Palaeodictyoptera, не зная о сильно специализированном ротовом аппарате последних). Представление о примитивности Anisoptera связано с тем, что по габитусу они сходны с некоторыми Palaeodictyoptera (рис. 7.4A) и Meganisoptera (рис. 7.3.5A): тело более массивное, чем у Zygoptera, крылья широкие, нестебельчатые, с густым жилкованием, задние крылья проксимально расширены. Названия палеозойских таксонов Meganisoptera и Protanisoptera связаны именно с этими гипотезами. Однако во всем остальном, кроме поверхностного сходства, Anisoptera не имеют общих признаков с названными вымершими насекомыми; в частности, ширина и густое жилкование крыльев в этих группах не связаны с одинаковым развитием каких-либо гомологичных жилок.
Была высказана гипотеза о независимом происхождении Zygoptera и Anisoptera от разных Meganisoptera (Tillyard 1917 и др.). Эта гипотеза основывалась на том, что у Anisoptera жилка iRS, в отличие от Zygoptera, снабжается трахеей не от того же ствола, что прочие ветви RS, а от трахеи, проходящей внутри R1 – так что эта трахея идет по поперечным жилкам и пересекает на своем пути RSa1 и RSa2. Полагая, что жилки насекомых возникли из трахей, исследователи делали вывод о том, что у Anisoptera жилка RA разветвляется, и ее задняя ветвь пересекает RSa1 и RSa2. Отсюда предполагали, что Anisoptera произошли от тех из Meganisoptera, у которых было описано ветвление RA (в действительности RA у всех Meganisoptera, как и у прочих Odonatoptera, не ветвится).
Была высказана также гипотеза о голофилии палеозойско-триасовой группы Odonatoptera, для которой на основании этого был установлен подотряд Meganeurina (Притыкина 1980) или отряд Meganeurida (Притыкина 1989):

При этом предполагают независимое возникновение очень сходных крыльев у Protozygoptera и Zygoptera. Однако для Meganeurida не указано ни одной надежно установленной апоморфии.
Предложена кладистическая классификация стрекоз (Bechly 1996), основанная на предположении о примитивности жилкования крыльев у Geroptera [см. Odonata (6) и ниже, Палеозойско-триасовые стрекозы]:

В этой классификации таксон Neodonatoptera Bechly 1996, объединяющий всех Odonatoptera кроме Geroptera, характеризуется суженными крыльями и тем, что MA отходит от RS, утратив связь с жилкой, считающейся исходным стволом M (обозначаемой нами как CuA – рис. 7.3.5C). Таксон Euodonatoptera Bechly et al. 2001 характеризуется отсутствием переднегрудных крыловидных параноталий и тем, что вместо архедиктия на крыльях имеется сеть из поперечных жилок; эти признаки считаются аутапоморфиями на основании спорных представлений о том, что переднегрудные параноты гомологичны крыльям [см. Pterygota (4D)] и что архедиктий первичен [см. Pterygota (4Ad)]. Таксон Odonatoclada Bechly et al. 2001, включающий всех Odonatoptera кроме Meganisoptera s.l., характеризуется тем, что жилки RA и RS слиты в основании. Таксон Nodialata Bechly 1996, включающий всех Odonatoptera кроме Meganisoptera s.l. и Campyloptera/fg, характеризуется наличием узелка (хотя у некоторых Protozygoptera и Protanisoptera узелок неясно выражен, без узелковой и подузелковой жилок) и утратой ветвления жилки MA; предполагают, что именно в этом таксоне появился вторичный копулятивный аппарат [см. Odonata (8)], тогда как у Meganisoptera он отсутствовал (Bechly 1998, публикация в Internet). Таксон Discoidalia Bechly 1996 {syn.circ.: Pandiscoidalia Nel et al. 2001}, объединяющий Triadophlebioptera, Protozygoptera и Neodonata, характеризуется прежде всего наличием поперечной жилки, ограничивающей дистально дискоидальную ячейку, тогда как проксимально эта ячейка может быть замкнутой или не замкнутой (хотя у некоторых Protozygoptera данный признак не выражен). Таксон Stigmoptera Bechly 1996, объединяющий Protozygoptera и Neodonata, характеризуется прежде всего наличием птеростигмы, ограниченной жилками, и не гомологичной птеростигме Protanisoptera.
Возможно, эта классификация в наибольшей степени соответствует филогении стрекоз. Однако в связи с тем, что многие важные признаки вымерших стрекоз неясны, в данном издании Odonatoptera (или Odonata s.l.) делятся на 5 таксонов равного ранга – вымершие Meganisoptera, Geroptera, Triadophlebioptera, Protozygoptera и Protanisoptera (см. Палеозойско-триасовые стрекозы) и представленные в современной фауне Neodonata (или Odonata s.str.).
Ниже рассмотрим палеозойско-триасовых стрекоз и Neodonata (стр. 0000).
Odonatoptera, или Odonata s.l. (или Libellula/fg1)
![]() 1.1 †Палеозйско-триасовые стрекозы
1.1 †Палеозйско-триасовые стрекозы
![]() 1.2 Neodonata, или Odonata s.str. (или Libellula/fg2)
1.2 Neodonata, или Odonata s.str. (или Libellula/fg2)
![]() 1.2.1 Zygoptera (или
Calepteryx/f1=Lestes/g1) – равнокрылые стрекозы
1.2.1 Zygoptera (или
Calepteryx/f1=Lestes/g1) – равнокрылые стрекозы
![]() 1.2.2 Anisopteroides (или Libellula/fg3)
1.2.2 Anisopteroides (или Libellula/fg3)
![]() 1.2.2.1 Anisoptera (или Libellula/fg4) – разнокрылые стрекозы
1.2.2.1 Anisoptera (или Libellula/fg4) – разнокрылые стрекозы
![]() 1.2.2.2 Epiophlebioptera (или
Epiophlebia/fg1)
1.2.2.2 Epiophlebioptera (или
Epiophlebia/fg1)
![]() 1.2.2.3 Вымершие группы, относящиеся к Anisopteroides
1.2.2.3 Вымершие группы, относящиеся к Anisopteroides