КЛАДОЭНДЕЗИС НАСЕКОМЫХ

![]()
|
КЛАДОЭНДЕЗИС НАСЕКОМЫХ |
|||||||
|
|
|
fg | |||||
X-1.3.1. Hymenoptera Linnaeus 1758 – перепончатокрылые
(Somobialia![]() Eucaryota
Eucaryota![]() Metazoa
Metazoa![]() Monostomata
Monostomata![]() Haemataria
Haemataria![]() Peritoneata
Peritoneata![]() Metameria
Metameria![]()
Gnathopoda![]() Euarthropoda
Euarthropoda![]() Mandibulata
Mandibulata![]() Atelocerata
Atelocerata![]() Hexapoda
Hexapoda![]() Amyocerata
Amyocerata![]()
Pterygota![]() Metapterygota
Metapterygota![]() Neoptera
Neoptera![]() Eumetabola
Eumetabola![]() Metabola
Metabola![]() Mecopteriformia
Mecopteriformia![]() Hymenoptera)
Hymenoptera)
(рис. 10.7.1–10.7.4)
(рис. 10.7.1,
10.7.2, 10.7.3,
10.7.4)
|
Названия, однозначно соответствующие по циркумскрипции: — Hymenoptera Linnaeus 1758; — Aculeata Scopoli 1763; — Piezata Fabricius 1793; — Phleboptera Clairville 1798; — Hymenopteria Rafinesque 1815; — Solenognatha Spinola 1850; — Metabola-Polynephria Brauer 1885 (non Menognatha-Polynephria Brauer 1885); — Lambentia Haeckel 1889; — Hymenopteroidea Handlirsch 1903; — Hymenopterida Pearce 1936; — Panhymenoptera Crampton 1938. По циркумскрипции также соответствует таксонам под типифицированными названиями, образованными от родового названия Vespa: Vespoides, Vespida, Vespidea. Иерархическое название: Vespa/f1=Cynips/g1 [f:1781(ord.6); g:L.1758(gen.213)] (incl. Xyela). |
|
Ранг. Обычно отряд, иногда одновременно с этим надотряд.
Аутапоморфии Hymenoptera
(1) Увеличение числа члеников щупиков. У имаго число члеников щупиков увеличено на один: максиллярный щупик [исходно 5-члениковый – см. Hexapoda (2c)] обычно имеет 6 члеников; лабиальный щупик [исходно 3-члениковый – см. Hexapoda (3c)] обычно имеет 4 членика. В некоторых таксонах произошло вторичное уменьшение числа члеников максиллярных и/или лабиальных щупиков, так что число члеников максиллярного щупика бывает от 1 до 6, лабиального щупика – от 1 до 4. Иногда щупик выглядит состоящим из бóльшего числа члеников благодаря дополнительному расчленению (рис. 10.7.1).
В отличие от имаго, у личинки сохраняется исходное для Hexapoda число члеников: максиллярный щупик имеет 5 или меньшее число члеников, иногда утрачен; лабиальный щупик имеет 3 членика, у Euhymenoptera одночлениковый (Расницын 1969).
(2) Модификация переднегруди. У имаго переднегрудь имеет характерное строение (рис. 10.7.1): латеральные цервикальные склериты [см. Pterygota (2)] неподвижно слиты с плейритами переднегруди; вся плейро-стернальная часть переднегруди (несущая передние ноги) подвижно сочленена с тергитом переднегруди и со среднегрудью [увеличенной – см. (5a)]. Тергит переднегруди может сохранять крупный размер и подвижное сочленение со среднегрудью (рис. 10.7.1), либо укорочен и слит со среднегрудью; в обоих случаях слитная плейро-стернально-шейная область переднегруди сохраняет подвижность по отношению к среднегруди.
(3) Модификация бедер. У имаго небольшой проксимальный участок бедра отделен от остальной части бедра кольцевым швом и выглядит как второй вертлуг; этот участок называют вертлужок (trochantellus), или префемур (praefemur) (рис. 10.7.1). Внешне кажется, что нога имеет «двучлениковый» вертлуг, тогда как истинный вертлуг здесь одночлениковый [как и у других насекомых – см. Hexapoda (5)]. Такой «двойной» вертлуг выражен на всех или на некоторых ногах многих Symphyta и Parasitica, но в некоторых случаях утрачен; в частности, он отсутствует у всех Monotrocha (см. «Классификации» I и II).
(4) Модификация шпоры передней ноги. У имаго на передней ноге имеется особый аппарат для чистки антенн – стригил (strigilis), что выражается в следующем (рис. 10.7.1). Передняя шпора [одна из двух вершинных шпор голени – см. Neoptera (4)] видоизменена: обычно слегка изогнута и снабжена коротким предвершинным или вершинным отростком, направленным к лапке; между этим отростком и основанием имеется продольная мембранозную каемка или гребень – велум (velum). Первый членик лапки в этом месте часто имеет более или менее выраженную выемку с щетинковидными отростками. Вторая (задняя) шпора передней голени может сохраняться, оставаясь неспециализированной, либо утрачена (у Unicalcarida – см. «Классификации» III).
У некоторых Strophandria обе шпоры передней голени имеют неспециализированное строение; вероятно, это вторичная утрата исходной для Hymenoptera специализации. В редких случаях шпоры передней голени утрачены.
(5) Модификация крыльев. Крыловой аппарат имеет ярко выраженную переднемоторность [см. (5a)]; жилкование крыльев модифицировано [см. (5b)]; задние крылья сцепляются с передними и движутся пассивно [см. (5c)].
У большинства перепончатокрылых крылья развиты; иногда крылья развиты только у самцов, а у самок вестигиальные или отсутствуют; реже крылья отсутствуют у обоих полов; у муравьев (Sodales, или Formica/fg) самки бесплодной касты бескрылые, тогда как самки плодовитой касты и самцы крылатые, но самки сбрасывают крылья после оплодотворения.
(5 a) Переднемоторность. Среднегрудь увеличена, заднегрудь уменьшена. Крылья имеют типичную для переднемоторных насекомых форму [см. Pterygota (4F)]: переднее крыло треугольное, с выраженным торнусом; заднее крыло всегда значительно меньше переднего, его костальный (передний) край равен по длине базиторнальному (заднему) краю переднего крыла; так что когда крылья сцеплены, задний край заднего крыла выглядит продолжением торноапикального (дистального) края переднего крыла. У всех перепончатокрылых торнус переднего крыла находится в дистальной части, т.е. базиторнальный край длиннее торноапикального. Переднее и заднее крыло сцепляются с помощью крючковидных щетинок, расположенных на переднем края заднего крыла и зацепляющихся за подогнутый базиторнальный край переднего крыла [см. (5с)]. Несмотря на то, что задние крылья всегда уменьшены и движутся пассивно, они почти никогда не утрачиваются, так что перехода к морфологической двукрылости не происходит (в отличие от многих других переднемоторных насекомых); лишь у одного вымершего вида Dipteromma paradoxa Rasnitsyn, Sidorchuk, Zhang & Zhang 2019 задние крылья полностью утрачены при наличии развитых передних крыльев.
(5 b) Жилкование крыльев (рис. 10.7.1, 10.7.2A–B). Имеется сравнительно небольшое число крепких постоянных жилок, соединенных под разными углами и образующими крупные многоугольные ячейки; при этом бывает трудно понять, какие из сторон этих ячеек образованы отрезками исходных продольных жилок, а какие – поперечными жилками.
На переднем крыле жилки Sc и RA более или менее сближены с C (идущей по переднему краю крыла); RA сливается с C несколько дистальнее середины переднего края крыла; в месте этого слияния имеется утолщение – птеростигма (pterostigma). Лишь в редких случаях птеростигма вторично утрачена. Жилка, идущая по переднему краю крыла дистальнее птеростигмы называется метакарп (metacarpus) (ее можно интерпретировать как продолжение C, либо как продолжение RA, либо как результат слияния C и RA). Жилка RS имеет не более двух ветвей и впадает в идущий по переднему краю крыла метакарп (две ветви RS имеются только у Archihymenoptera – рис. 10.7.1, у остальных RS не ветвится – рис. 10.7.2A). Дистальнее места слияния с RS метакрп не продолжается, так что весь остальной край крыла (вершинный и задний) не имеет краевой жилки. Основания жилок RS, M и Cu связаны между собой косыми жилками так, что от основания крыла между RA и Pcu идет единственная продольная жилка (считающаяся либо Cu, либо результатом слияния M+Cu); иногда она отходит не от основания крыла, а от RA. Жилки M и Cu1 не ветвятся и, если их дистальные части развиты, то они оканчиваются у наружного (торноапикального) края крыла. От Cu назад отходит короткая жилка Cu2, своей вершиной она сливается с вершиной Pcu; это слияние, пересекающее клавальный желоб, находится в дистальной части крыла вблизи заднего (базиторнального) края, около дистального конца подгиба заднего края, обеспечивающего сцепку крыльев [см. (5c)]. Ваннальная жилка (V), если имеется, тянется вдоль заднего края крыла и впадает в Pcu.
В случаях крайней редукции жилкования (независимо произошедшей в нескольких неродственных друг другу таксонах) на переднем крыле имеется лишь RA, птеростигма (или краевая жилка на ее месте) и отходящий от нее назад-дистально короткий отрезок RS.
Заднее крыло [в полете сцепляющееся с передним – см. (5c)] по жилкованию более или менее сходно с передним, но имеет определенные отличия. Инверталярная область заднего крыла [переворачивающаяся при складывании – см. Neoptera (1a)] бывает развита в виде выступающей лопасти, называемую «югальной», или «анальной». Исходно инверталярная лопасть несет продольную жилку (рис. 10.7.1, 10.7.2B). У многих Apocrita инверталярная лопасть утрачена.
Нигмы [см. Metabola (5)] имеются только у Symphyta (Мартынов 1924). На переднем крыле бывает по одной нигме в следующих ячейках (рис. 10.7.2A): в проксимальной ячейке между RA и Cu; в первой ячейке между RS и M; во второй ячейке между RS и M; во второй ячейке между M и Cu (отсутствует на рис. 10.7.2A). На заднем крыле бывает по одной нигме в следующих ячейках: в единственной ячейке между RS и M; в единственной ячейке между Pcu и V1 (рис. 10.7.2B).
(5 c) Сцепка крыльев (рис. 10.7.1, 10.7.2A–B). Заднее крыло сцепляется с передним с помощью крючковидных щетинок (hamuli), которые образуют правильный продольный ряд на дорсальной стороне костального края заднего крыла на участке, гомологичном птеростигме [см. (5b)]. Эти щетинки зацепляются за подогнутый вентрально базиторнальный край переднего крыла, который тянется до места слияния Cu2 и Pcu и укреплен этим слиянием. Кроме сцепочных щетинок, на переднем крае заднего крыла бывают развиты еще одна или две более проксимальные группы крючковидных щетинок, не служащих для сцепки (рис. 10.7.1, 10.7.2B). Сходный сцепочный аппарат имеется у некоторых Parametabola (см. Указатель признаков [2.152]).
(6) Складывание крыльев. В покое крылья складываются [см. Neoptera (1c)] обычно плоско на спине. Исходно имеется уникальный аппарат для фиксации сложенных таким образом крыльев: на заднеспинке находятся ценхри (cenchri) – это пара мембранозных овальных участков или лопастинок, возвышающихся над поверхностью заднеспинки; в задне-проксимальной части переднего крыла имеется участок, густо покрытый шиповидными микротрихиями, направленными дистально; ваннальная жилка образует изгиб, окаймляя этот участок спереди (рис. 10.7.1); когда крыло укладывается на спину, этот шероховатый участок смыкается с ценхром. У Cephus/fg и у всех Apocrita (составляющих подавляющее большинство Hymenoptera) ценхри утрачены.
(7) Модификация склеритов брюшка. У имаго тергиты прегенитальных сегментов брюшка занимают не только дорсальные, но и латеральные поверхности сегментов; их латеральные лопасти, называемые эпиплеврами (epipleura), направлены вентрально и облегают латеральные области стернитов (кроме первого сегмента брюшка, стернит которого отсутствует); при этом дыхальца [на I–VIII уромерах или на части из них – см. Amyocerata (2)] находятся на склеротизованных тергитах (рис. 10.7.1). То же у некоторых других насекомых.
(8) Гениталии самца. Гениталии самца [см. Metabola (6)] у всех Hymenoptera имеют единый план строения (рис. 10.1.4B и 10.7.2C–D) (Snodgrass 1957, Schulmeister 2001). Основание генитального тегмена сужено и соединено с туловищем посредством эластичной мембраны. Задний край IX стернита брюшка образует пластину (называемую гипандрий, или гипопигий), которая выступает назад от места прикрепления генитального тегмена и закрывает его в покое. Благодаря разнонаправленным мышцам, идущим от основания генитального тегмена к IX стерниту брюшка и к гипандрию, весь фаллический комплекс может как единое целое выдвигаться, отклоняться или переворачиваться на 180º вокруг продольной оси. Самая базальная часть генитального тегмена, называемая купула (cupula), обычно имеет вид обособленного склеротизованного кольца, но иногда она частично или полностью слита с остальным тегменом или редуцирована. Вместо одной пары парамеров в составе тегмена имеются две пары хватательных придатков – пара более длинных латеральных гонофорцепсов и пара более коротких медиальных вольселл. Каждый гонофорцепс, или латимер, исходно состоит из двух подвижных члеников – проксимального гоностипеса и дистального телопарамера. Гоностипес (gonostipes) представляет собой склеротизованную латеральную стенку генитального тегмена; на дорсальной стороне тегмена левый и правый гоностипесы соприкасаются или слиты между собой по средней линии; на вентральной стороне тегмена гоностипесы соприкасаются лишь в самой проксимальной точке. К вершине каждого гоностипеса причленен подвижный придаток – телопарамер, или харпаго. У многих Unicalcarida гонофорцепсы цельные, не расчлененные на гоностипес и харпаго. На вентральной стороне тегмена между гоностипесами расположены вольселлы (volsellae) – парные склериты, выступающие дистально в виде коротких склеротизованных отростков – пары подвижных дигитусов (digiti) и пары расположенных по бокам от них кусписов (cuspides); дигитус и куспис каждой вольселлы образуют зажим. Такие щипцевидные вольселлы являются новообразованием; они имеются у большинства перепончатокрылых, но у пчелиных редуцированы или утрачены. Эдеагус выступает дистально из генитального тегмена дорсальнее вольселл, между гонофорцепсами. Латеральные стенки эдеагуса укреплены парой продольных склеритов – пенисвальв (penisvalvae); в дистальной части пенисвальвы обычно на большем или меньшем протяжении обособлены в виде пары склеротизованных выростов. От основания каждой пенисвальвы вглубь генитального тегмена уходит длинная склеротизованная аподема. Вторичное половое отверстие открывается наружу либо на мембранозной вентральной поверхности генитального тегмена в основании эдеагуса, либо на самом эдеагусе между пенисвальвами, либо (у пчелиных) на вершине особого непарного трубчатого отростка, выступающего между парой обособленных пенисвальв. У разных перепончатокрылых генитальный аппарат имеет те или иные мышцы, идущие в следующих направлениях: от купулы к основаниям гоностипесов; от разных частей гоностипесов к пенисвальвам и их аподемам; от разных частей вольселл к аподемам пенисвальв; от разных частей гоностипесов к разным частям вольселл; между разными частями вольселл; от гоностипеса к харпаго; кроме того, иногда имеются мышцы, соединяющие левый и правый гоностипесы или аподемы пенисвальв. Таким образом, здесь, как и у прочих Mecopteriformia и Eleuterata, эдеагус выдвигается мышцами, крепящимися внутри парамеров генитального тегмена – т.е. внутри гонофорцепсов и вольселл. При копуляции гонофорцепсы охватывают VII стернит брюшка самки; вольселлы своими дигитусами и кусписами зажимают либо задний край того же стернита, либо сочленовную мембрану у генитального отверстия; пенисвальвы, благодаря своим аподемам, выдвигаются и проникают в половое отверстие самки.
(9) Модификация личиночных глаз. У личинки каждый латеральный глаз представляет собой единый нефасеточный глаз, образованный слиянием омматидиев фасеточного глаза [см. Mandibulata (4)]; он имеет единую выпуклую кутикулярную линзу, под ней находится множество рабдомов, каждый из которых представляет собой рабдом одного исходного омматидия и, как в омматидиях большинства насекомых, состоит из 8 клеток; кристаллические конусы утрачены (Meyer-Rochow 1973) [тогда как у большинства других личинок насекомых с полным превращением фасеточный глаз состоит из 6–7 разрозненных стемм – см. Metabola (1Ca)]. Глаза имеются у личинок многих Symphyta (рис. 10.7.3A), но у Urocerus/f1=Sirex/g1 и Euhymenoptera (составляющих большинство видов перепончатокрылых) личиночные глаза полностью утрачены в связи со скрытым образом жизни (рис. 10.7.4A). Неуникальная апоморфия; сходные сложные нефасеточные глаза имеются у некоторых других членистоногих.
(10) Гапло-диплоидное определение пола и арренотокический партеногенез. Из оплодотворенных яиц развиваются диплоидные самки, а из неоплодотворенных яиц – гаплоидные самцы [см. Metazoa (2)]. Соответственно, только у самок оогенез сопровождается мейозом, тогда как у самцов сперматогенез происходит без мейоза. Благодаря гапло-диплоидному определению пола степень генетического сходства между родными сестрами, получившими половину своих генов от одного и того же гаплоидного самца, составляет 3/4 (тогда как между матерью и дочерью она лишь 1/2); так что естественный отбор может поддерживать такие формы поведения, при которых самка выращивает своих сестер в ущерб собственным дочерям. Благодаря этому в некоторых таксонах жалящих перепончатокрылых, у которых самки выкармливают личинок в гнезде, независимо несколько раз возникали одинаковые формы эусоциальности (см. «Классификации» III: Monotrocha).
Помимо Hymenoptera, гапло-диплоидное определение пола обнаружено у изученных в этом отношении Thysanoptera, у отдельных представителей Eleuterata, Arthroidignatha, Monomerosomata и Porifera (White 1984); функциональная гаплодиплоидия свойственна многим Gallinsecta, среди которых есть и вполне гаплодиплоидные представители (Gullan & Cook 2007).
Признаки Hymenoptera неясного филогенетического значения
(11) Строение антенн. У имаго число члеников флагеллума антенны [см. Metabola (1Cbβ)] бывает неопределенным или определенным, от одного до нескольких десятков (cм. Указатель признаков [1.27]: табл. B). У Archihymenoptera первый членик флагеллума (т.е. третий членик антенны) значительно длиннее и толще остальных (рис. 10.7.1); если исходить из предположения, что Archihymenoptera является предковым таксоном для остальных перепончатокрылых (см. «Классификации» III), то такое модифицированное строение антенны может быть исходным для Hymenoptera (Расницын 1969). Одночлениковый флагеллум (т.е. 3-члениковая антенна) имеется у Strophandria-Arge/fg; в этом случае единственный членик флагеллума длинный, как первый членик у Archihymenoptera или многочлениковый флагеллум у других перепончатокрылых.
(12) Срединные шпоры на голени. У имаго голени могут иметь, помимо пары апикально-внутренних гиподермосодержащих первичных шпор [см. Neoptera (4) и Metabola 1Cc)], расположенные сходными парами срединно-внутренние гиподермосодержащие шпоры (называемые также предвершинными, или надвершинными шпорами) (рис. 10.7.1). То же у ручейников и бабочек [ср.: Amphiesmenoptera (7)]. Максимальный набор шпор у Hymenoptera следующий: на каждой голени может быть до 2 апикально-внутренних шпор [об их модификации на передней ноге – см. (4)]; на передней голени кроме этой пары апикально-внутренних шпор, может быть одна срединно-внутренняя шпора (у Acantholyda/g в составе Pamphilius/fg1); на средней голени, кроме пары апикально-внутренних шпор, может быть до 3 срединно-внутренних шпор – пара шпор проксимальнее апикально-внутренних и одна шпора еще проксимальнее (т.е. всего до 5 шпор); на задней голени, кроме пары апикально-внутренних шпор, может быть до 4 срединно-внутренних шпор – пара шпор проксимальнее апикально-внутренних и пара шпор еще проксимальнее (т.е. всего до 6 шпор). Срединно-внутренние шпоры развиты лишь у некоторых представителей плезиоморфона Symphyta – Archihymenoptera (рис. 10.7.1), Pamphilius/fg, Strophandria-Arge/fg и Cephus/fg. У большинства перепончатокрылых срединно-внутренние шпоры утрачены.
(13) Увеличение числа мальпигиевых сосудов. У имаго мальпигиевы сосуды многочисленные, их бывает от 8 до 125 [в отличие от всех прочих Metabola, имеющих не более 8 сосудов – см. Eumetabola (5)]. В отличие от имаго, у личинки число мальпигиевых сосудов не увеличено, обычно от 4 до 8 (т.е. от 2 до 4 пар). У многих эндопаразитических личинок мальпигиевы сосуды утрачены.
С этим признаком связано название Polynephria Brauer 1885. Такое же неопределенно большое число мальпигиевых сосудов считается исходным для Pterygota (поскольку присуще Ephemeroptera, Odonata, Idioprothoraca и Rhipineoptera – см. Указатель признаков [4.7]: табл. F), тогда как у Eumetabola число мальпигиевых сосудов сокращено и стабилизировано. В связи с этим Брауер (Brauer 1885) противопоставлял перепончатокрылых (Hymenoptera, или «Metabola polynephria») всем прочим насекомым с полным превращением, которых объединял в таксон под названием «Metabola oligonephria». Судя по тому, что у личинок и части имаго перепончатокрылых имеются лишь 2–4 пары мальпигиевых сосудов (как у других насекомым с полным превращением), большое число мальпигиевых сосудов у части имаго перепончатокрылых является вторичным.
Плезиоморфии и изменчивые признаки Hymenoptera
(14) Грызущие мандибулы. У имаго мандибулы [см. Metapterygota (2)] всегда хорошо развиты и всегда сохраняют исходную грызущую функцию.
Другие части ротового аппарата – максиллы, нижняя губа и гипофаринкс – также могут также сохранять строение, свойственное исходному грызущему ротовому аппарату [см. Hexapoda (1)], либо преобразованы и адаптированный для питания нектаром цветков. В последнем случае складывающиеся и выдвигающиеся части лабио-максиллярного комплекса [см. Mecopteriformia (2)] сильно удлинены, так что образуют лижуще-сосущий хоботок, способный выдвигаться и проникать в нектарники цветков. Такой хоботок независимо возник в разных таксонах перепончатокрылых.
(15) Первичный яйцеклад. У самки обычно сохраняется первичный яйцеклад, состоящий из 3 пар створок и пары гонангулов [см. Amyocerata (7)]; при этом первые и вторые створки (т.е. пара VIII кинетапофизов и слитые между собой IX кинетапофизы) сцеплены олистетерами и образуют режущий или колющий яйцеклад, а третьи створки (придатки IX уромера, считающиеся стилигерами и/или стерностилями) образуют ножны характерного строения [см. Eumetabola (1)] (рис. 10.7.1).
Среди Metabola перепончатокрылые сохраняет наиболее полное строение первичного яйцеклада (подвижно сочлененные гонангулы, стилигеры IX уромера и две пары кинетапофизов), тогда как у всех прочих Metabola яйцеклад в той или иной мере упрощен или утрачен [см. Eumetabola (1)]. Следует обратить внимание на очень большую консервативность в сохранении первичного яйцеклада у Hymenoptera: у представителей с самыми различными способами откладки яиц яйцеклады по-разному модифицированы и по-разному функционируют, но сохраняют все элементы исходного плана строения, лишь в редких случаях подвергаются редукции.
Вероятно, исходным для яйцеклада Hymenoptera является наличие на кинетапофизах поперечных или косых гребней, называемых «аннули»; на вентральном крае первых створок (VIII кинетапофизов) и/или на дорсальном крае вторых сворок (IX кинетапофизов) эти гребни могут особенно сильно выступать, придавая краю яйцеклада пиловидную форму. У представителей плезиоморфона Symphyta, личинки которых питаются травами и листьями (Strophandria, Pamphilius/fg и Cephus/fg), яйцеклад приспособлен для пропиливания ткани растения: он недлинный, с пиловидным вентральным краем первых створок; в связи с такой формой и функцией яйцеклада этих насекомых называют пилильщиками.
У многих других перепончатокрылых (Xyela/fg, Urocerus/f=Sirex/g, многие Euhymenoptera) яйцеклад прямой, удлиненный, приспособлен для прокалывания; косые гребни на его сворках бывают развиты или отсутствуют. У Euhymenoptera колющий яйцеклад, помимо функции проведения яиц, может использоваться для проведения ядовитого секрета; с помощью такого яйцеклада-жала самка парализует насекомое и откладывает на него яйца. У Euhymenoptera-Monotrocha яйцеклад утратил функцию проведения яиц и сохранил только функцию жала. Лишь у немногих перепончатокрылых яйцеклад утрачен.
Размеры. Длина тела 0,2–60 мм.
Геологический возраст. Триас – ныне (рис. 2.2). Наиболее древними известными перепончатокрылыми являются представители современного таксона Archihymenoptera из раннего триаса Средней Азии (Расницын 1969; Rasnitsyn & Quicke 2002).
Состав и распространение. По разным оценкам от 100 000 до 200 000 видов и более; всесветно.
Систематическое положение Hymenoptera. Помимо принятого здесь отнесения Hymenoptera непосредственно к Mecopteriformia, предлагались следующие классификации.
I. Отнесение Hymenoptera непосредственно к Metabola и противопоставление их Aparaglossata (см. Metabola: «Классификации» II).
II. Объединение Hymenoptera и Elytrophora в таксон Necromorpha (см. Elytrophora: «Систематическое положение» II).
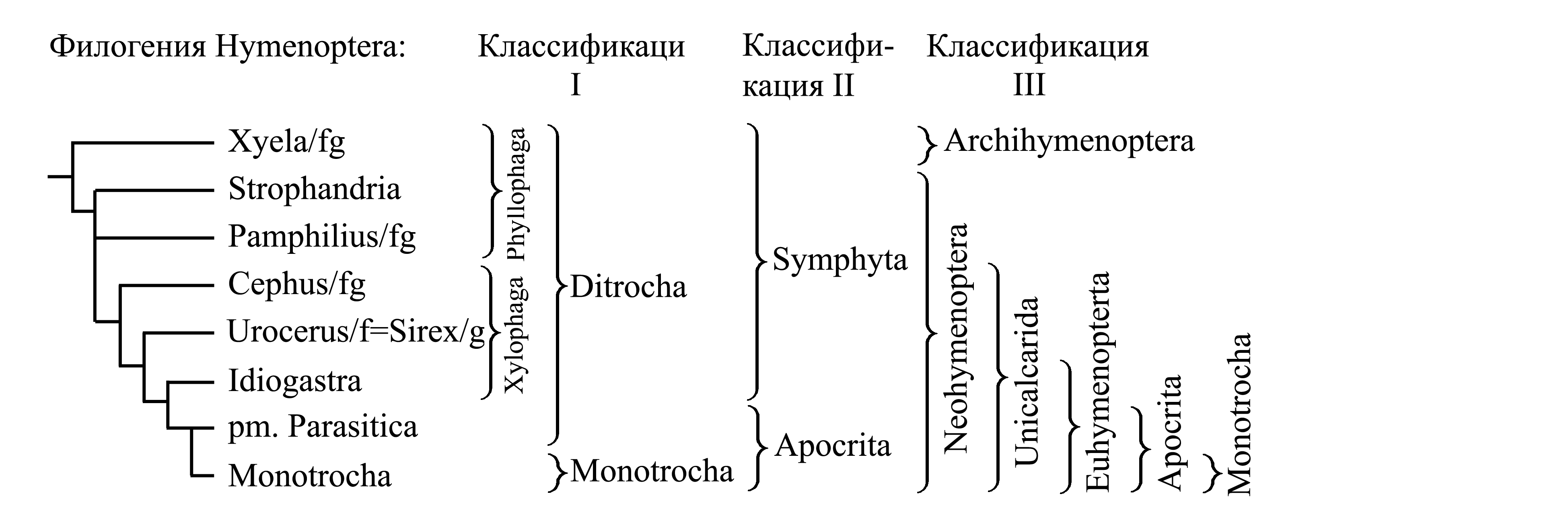
Классификации Hymenoptera
I. Деление Hymenoptera на Ditrocha и Monotrocha. Исходно перепончатокрылых делили на имеющих яйцеклад и имеющих жало (Latreille 1802–1825, Leach 1815 и др.) [см. Hymenoptera (15)]; таксон, характеризовавшийся наличием яйцеклада, называли Terebrantes Latreille 1802b {syn.circ.: Terebrantia Latreille 1810}, а таксон, характеризовавшийся наличием жала – Aculeati Latreille 1802b {= «Aculeata»: Latreille 1807 (non Scopoli 1763)}. При этом ос-блестянок (Chrysis/fg), имеющих вторичный яйцеклад, либо относили к Terebrantes, либо выделяли в отдельный таксон Tubuliferi Latreille 1804c, но не помещали в состав жалящих. По современным представлениям осы-блестянки должны объединяться с жалящими; в этом случае классификация выглядит следующим образом.
1. Ditrocha Hartig 1837.
1.1. Symphyta (см. II) {= Phyllophaga Hartig 1837 + Xylophaga Hartig 1837}.
1.2. Parasitica Hartig 1837.
2. Monotrocha Hartig 1837 (non Monotrocha Ehrenberg 1830) (= Aculeati Latreille 1802b + Tubuliferi Latreille 1804c).
В этой классификации таксон Ditrocha (близкий по составу к Terebrantes) характеризуется «двучлениковыми верлугами» [т.е. наличием вертлужка – см. Hymenoptera (3)], тогда как у представителей таксона Monotrocha (близкого по составу к Aculeati) ноги имеют обычное строение, без вертлужка. Если основываться только на этом признаке, можно было бы предположить, что наличие вертлужка является аутапоморфией, а таксон Ditrocha является голофилетическим; однако это противоречит представлению о голофилии Apocrita (см. III). По всей видимости, отсутствие у Monotrocha вертлужка является не плезиоморфией, а результатом вторичной утраты. В настоящее время таксон Ditrocha не признают, считая его парафилетическим, а таксоны Monotrocha и Parasitica рассматривают как подчиненные в составе Apocrita (см. II и III).
II. Деление Hymenoptera на Symphyta и Apocrita. Если в предыдущей классификации (см. I) часть перепончатокрылых со стебельчатым брюшком помещена в плезиоморфон Ditrocha (в качестве подчиненного таксона Parasitica), а часть составляет таксон Monotrocha, то здесь они объединены в голофилетический таксон Apocrita. Прочих перепончатокрылых обычно объединяют в плезиоморфон Symphyta. Такая классификация остается широко признанной до настоящего времени.
1. Symphyta Gerstaecker 1867 {synn.circ: Ventrosessiles Latreille 1802b = Sessiliventres Latreille 1804c = Serricaudia Dumeril 1805 = Uroprista Dumeril 1805 = Phytiphaga Latreille 1807 = Pristuria Rafinesque 1815 = Securifera Latreille 1817 = Phytophaga Billberg 1820 (non Phytophaga Dumeril 1805) = Serrifera Berthold 1827 = Phytospheces Leunis 1860 = Chalastogastra Konov 1897 = Prohymenoptera Crampton 1916a}.
2. Apocrita Gerstaecker 1867.
2.1. Parasitica Hartig 1837.
2.2. Monotrocha Hartig 1837 (non Monotrocha Ehrenberg 1830).
III. Деление Hymenoptera на Archihymenoptera и Neohymenoptera. Эта классификация отражает современные представления о филогении перепончатокрылых (Rasnitsyn & Quicke 2002):
1. Archihymenoptera Grimaldi & Engel 2005 {nom.hier.: Xyela/fg1 [f:1834; g:1819] (incl. Macroxyela)}. Отличается от Neohymenoptera следующими плезиоморфиями: (1) на переднем крыле жилка RS двуветвистая (рис. 10.7.1) [см. Hymenoptera (5b)]; (2) куколка с действующими мандибулами – «pupa dectica» (см. Указатель признаков [5.24]). Второй из этих признаков известен лишь по отдельным указаниям (Hinton 1971).
Мелкие, менее 5 мм. Яйцеклад длинный, выступает далеко за вершину брюшка. Личинки развиваются либо в мужских стробилах сосен, либо в почках и молодых побегах хвойных, либо на листьях вязовых и ореховых. Archihymenoptera являются наиболее древними среди перепончатокрылых, известны начиная с триаса. Включают около 50 современных видов, обитают только в Голарктике (преимущественно в Неарктике).
У имаго всех Archihymenoptera, как современных, так и ископаемых, флагеллум антенны имеет характерное строение, не встречающееся у других перепончатокрылых: его проксимальная часть представляет собой единый длинный толстый членик, а дистальная часть состоит из 9 или более члеников обычного размера (рис. 10.7.1). По мнению некоторых авторов, Archihymenoptera – это голофилетический таксон, аутапоморфией которого является особое строение антенн (Grimaldi & Engel 2005 и др.).
Согласно другой точке зрения, Archihymenoptera – это парафилетический таксон, предковый для Neohymenoptera. У куколки Archihymenoptera на увеличенном первом членике флагеллума обнаружены поперечные насечки, свидетельствующие о том, что этот членик образовался путем слияния нескольких проксимальных члеников; на основании этого выдвинуто предположение, что первый членик флагеллума Archihymenoptera сохранил наследственную информацию об утраченном расчленении, что проявляется на стадии куколки; в ходе дальнейшей эволюции это исходное расчленение могло восстановиться и на имагинальных антеннах у Neohymenoptera (Расницын 1969).
2. Neohymenoptera Grimaldi & Engel 2005 {nom.hier.: Vespa/f2=Cynips/g2 [f:1781; g:L.1758(gen.213)] (sine Xyela; incl. Tenthredo). По мнению одних авторов, это голофилетический таксон (Grimaldi & Engel 2005 и др.), по мнению других – полифилетический таксон, разные представители которого произошли от разных Archihymenoptera (Расницын 1969). Таксон Neohymenoptera характеризуется следующими аутапоморфиями.
(1) На переднем крыле развилок RS утрачен (рис. 10.7.2A) [см. Hymenoptera (5b)].
(2) Куколка без с неподвижными мандибулами [«pupa adectica» – см. Metabola (1Bb)], не передвигается в течение всего своего развития (см. Указатель признаков [5.24]). Обычно куколка находится в коконе, в этом случае имаго сбрасывает куколочную шкурку и расправляет крылья, оставаясь в коконе; выход имаго из кокона происходит только после склеротизации покровов, и имаго прогрызает кокон отвердевшими мандибулами [которые всегда развиты – см. Hymenoptera (14)]. Помимо Hymenoptera, такой же способ выхода из куколочного укрытия только у Elytrophora; у других насекомых с полным превращением расправление крыльев происходит вне укрытия [см. Metabola (1D)]. У большинства Neohymenoptera куколка свободная (pupa adectica exarata), но у некоторых Eulophus/f куколка склеенная (pupa conglutinata) (неуникальная апоморфия – см. Указатель признаков [5.24]) (Hinton 1946, 1971).
В состав Neohymenoptera входят Strophandria, Pamphilius/fg и Unicalcarida.
2.1. Strophandria Ross 1937 – пилильщики {nom.hier: Tenthredo/fg1 [f:1802; g:L.1758(gen.214)] (incl. Cimbex, Diprion, Arge)}. Голофилетический таксон. У имаго пронотум медиально сильно укорочен, с глубоко выемчатым задним краем. Гениталии самца [способные поворачиваться вокруг своей оси – см. Hymenoptera (8)] в покое у имаго и у куколки перевернуты на 180° (то же у имаго, но не у куколки Archihymenoptera-Xyela/fg2). Личинки, называемые ложногусеницами, похожи на гусениц бабочек: удлиненные, сегменты груди и брюшка сходного размера и строения; ноги развиты [см. Metabola (1Cc) и Mecopteriformia (4)]; второй и последующие сегменты брюшка несут по паре ложноножек, так что бывают развиты 8 или меньшее число пар ложноножек; ложноножки расположены на одной линии с грудными ногами (рис. 10.7.3A). Несмотря на то, что личинка сходна с гусеницей бабочки и имеет такие же короткие ноги, в процессе превращения личиночных ног в куколочные гиподерма ноги сначала сильно укорачивается, и лишь затем начинает расти (рис. 10.7.3C–D) [см. Metabola (1Ccδ-8); в отличие от Lepidoptera – см. Metabola (1C cδ-12)]. Личинки многих видов живут открыто на растениях, питаясь их зелеными частями; у некоторых видов личинки постоянно или временно обитают внутри растений или в галлах. Включает около 7000 видов, т.е. большую часть видов сидячебрюхих; распространены всесветно. Ископаемые известны начиная с юры.
2.2. Pamphilius/fg1 [f:1890; g:1802] (incl. Megalodontes) – паутинные пилильщики. Около 250 видов в Голарктике. Ископаемые известны начиная с юры.
2.3. Unicalcarida Schulmeister, Wheeler & Carpenter 2002 {nom.hier.: Vespa/f3=Cynips/g3 (sine Tenthredo; incl. Cephus, Sirex). Вероятно, голофилетический таксон, характеризуется следующими аутапоморфиями.
(1) У имаго на передней ноге задняя шпора утрачена, так что имеется только хорошо развитый стригил [видоизмененная передняя шпора – см. Hymenoptera (4)].
(2) Личинки ведут скрытый образ жизни; их ноги сильно уменьшены, нерасчлененные, без претарсуса (у Euhymenoptera совсем утрачены).
В состав Unicalcarida входят Apocrita и некоторые таксоны, традиционно относимые к плезиоморфону Symphyta – Cephus/fg и Urocerus/f=Sirex/g.
2.3.1. Cephus/fg1 [f:1834; g:1802] (incl. Hartigia) – стеблевые пилильщики. У имаго тело длинное и тонкое; пронотум удлинен; первый и второй сегменты брюшка разделены вдавлением, напоминающем стебелек у Apocrita (см. ниже). Ценхри и поле шипиков позади ваннальной жилки переднего крыла отсутствуют (то же у Apocrita), но ваннальная жилка сохраняется (в отличие от Apocrita). Личинки развиваются внутри стеблей растений: личинки Hartigia/fg1 [f:1914; g:1838] обитают в побегах и стеблях двудольных (розоцветных, ивовых, дубов); личинки Cephus/fg2 обитают в стеблях злаков. Около 100 видов. Ископаемые известны начиная с юры.
2.3.2. Urocerus/f1=Sirex/g1 [f:1802; g:1761] (incl. Syntexis, Xiphydria) – рогохвостовые. Личинки живут в древесине и питаются благодаря симбиозу с грибом, которым самка заражает дерево. В брюшке имаго самки имеется пара мешковидных органов – микангиев (mycangia), открывающихся протоками в основании яйцеклада. У личинки самки в складке между первым и вторым сегментами брюшка имеется пара углублений, выстланных железистым эпителием и служащих для культивирования гриба. При развитии имаго из личинки споры культивируемого гриба попадают в микангии. При откладке яиц самка выделяет придаточными железами яйцеклада (гомологичными ядовитым железам жалящих) секрет, который вместе со спорами гриба инъецируется в древесину. Разные виды рогохвостовых используют разные виды грибов, относящихся с аскомицетам и базидиомицетам (Gauld & Bolton 1988). Помимо рогохвостовых, микангии, служащие для переноса спор грибов, имеются у некоторых древоядных жуков, но они расположены в других частях тела и не гомологичны микангиям рогохвостов. Некоторые авторы не признают этот таксон, считая его парафилетическим, и рассматривают Urocerus/f=Sirex/g и Xiphydria/fg как независимые друг от друга как таксоны (Grimaldi & Engel 2005).
2.2.2.1. Anaxyela/f1=Syntexis/g1 [f:1925; g:1915] с единственным современным видом Syntexis libocedrii Rohwer 1915 в Калифорнии, личинки в древесине кипарисовых. Ископаемые известны из юры и мела.
2.3.2.2. Bomboptera MacLeay 1821 – рогохвосты {syn.circ.: Terebellifera Le Peletier 1827; nom.hier.: Urocerus/f2=Sirex/g2 [f:1802; g:1761]}. У имаго последний (десятый) сегмент брюшка с направленным назад роговидным выступом (с чем связано название Urocerus); яйцеклад сверловидный. Крупные, обычно более 15 мм. Личинки в древесине лиственных и хвойных деревьев. Около 100 видов. Ископаемые известны начиная с юры.
2.3.2.3. Xiphydria/fg1 [f:1817; g:1802]. Яйцеклад сохраняет пиловидную форму (в связи с чем в старых классификациях ксифидриид объединяли с пилильщиками). Личинки в древесине лиственных деревьев. Около 100 видов.
2.3.3. Euhymenoptera Engel & Grimaldi 2004 {nom.hier.: Vespa/f4=Cynips/g4 (sine Sirex; incl. Orussus)}. Голофилетический таксон, характеризуется следующими аутапоморфиями.
(1) Личинка исходно ведет паразитоидный образ жизни: питается на живом насекомом-хозяине (подобно паразиту), а в конце своего развития убивает и доедает его (подобно хищнику). Жертвами (или, как их часто называют, хозяевами) в большинстве случаев являются личинки и куколки насекомых с полным превращением, неподвижные или малоподвижные имаго некоторых Plantisuga и яйца различных насекомых и пауков. Паразитоидное развитие свойственно большинству видов Euhymenoptera. В наиболее примитивном случае самка при откладке яйца парализует хозяина уколом яйцеклада, а личика развивается на поверхности хозяина как эктопаразитоид. При этом иногда жертва остается парализованной на протяжении всего питания личинки. Иногда парализация временная, в таком случае личинка питается на жертве, ведущей активный образ жизни. Иногда парализации вообще не происходит, так что самка откладывает яйцо на активное насекомое. В некоторых таксонах Euhymenoptera-Apocrita личинки адаптировались к жизни внутри тела насекомого (эндопаразитоиды). У разных видов паразитоидов, в зависимости от соотношения размеров паразитоида и жертвы, на одной жертве может развиваться одна или много личинок. Личинки некоторых Euhymenoptera-Apocrita перешли от паразитоидного развития на насекомых-галлообразователях к самостоятельному галлообразованию, то есть являются фитофагами. Некоторые представители Euhymenoptera-Apocrita перешли от паразитоидного образа жизни к выкармливанию личинок в гнезде (см. ниже, Monotrocha).
(2) В связи с паразитоидным образом жизни личинки полностью утратили ноги [исходно вестигиальные – см. Unicalcarida]. У изученных представителей Euhymenoptera-Apocrita перед линькой личинки на куколку образующиеся под личиночной кутикулой гиподермальные зачатки куколочных ног вначале погружаются своими вершинами в акрогенные карманы (рис. 10.7.4A–C), и только позже, когда стенки карманов рассасываются, оказываются непосредственно под кутикулой [см. Metabola (1Ccδ-9)].
Таксон Euhymenoptera делится на Idiogastra и Apocrita.
2.3.3.1. Idiogastra Rohwer & Cushman 1917 – оруссиды {syn.circ.: Protoparasitica Engel 2005; nom.hier.: Orussus/fg1 [f:1834; g:1796]}. У имаго места прикрепления антенн смещены на вентральную сторону головы, под нависающий край лица; у самца антенна 11-члениковая, у самки 10-члениковая. У самки антенны и передние ноги приспособлены для распознавания места на стволе дерева, под которым находится личинка жука или рогохвоста. Яйцеклад очень длинный, не менее чем вдвое длиннее тела, в покое свернут внутри брюшка; приспособлен для откладки яиц в древесину. Личинки развиваются в ходах, проложенных в мертвой древесине другими насекомыми и могут питаться как эктопаразитоиды на личинках жуков и рогохвостов; они также могут питаться разлагающейся древесиной, подобно личинкам рогохвостов (Gauld & Bolton 1988). В отличие от Apocrita, у них сохраняется проход из средней кишки в заднюю, но кутикула задней кишки имеет складки, позволяющие ей растягиваться и откладывать дефекацию до предкуколочного состояния. Включает около 75 видов, распространенных во всех частях света; наибольшее число видов в южном полушарии (Vilhelmsen 2003). Разные авторы либо объединяют оруссид с Apocrita в таксон Euhymenoptera, либо объединяют оруссид с рогохвостами.
2.3.3.2. Apocrita Gerstaecker 1867 {syn.circ.: Petiolata Sharp 1894; nom.hier.: Vespa/f5=Cynips/g5 (sine Orussus). Голофилетический таксон, характеризуется следующими аутапоморфиями.
(1) У имаго и куколки тергит первого сегмента брюшка слит с грудью (стернит первого сегмента не выражен, как и у прочих Hymenoptera); основание второго сегмента брюшка суженное и очень подвижно сочленено с первым сегментом (рис. 10.7.4D). Таким образом кажущаяся граница между «грудью» и «брюшком» проходит по узкому подвижному сочленению между первым и вторым сегментами брюшка. Отдел тела, кажущийся «грудью» и образованный тремя грудными сегментами и истинным первым сегментом брюшка, называется мезосома (mesosoma); истинный первый сегмент брюшка, слитый с грудью и кажущийся ее частью, называется промежуточный сегмент, или проподеум (propodeum); сочленение между ним и истинным вторым сегментом брюшка называется стебелек (petiolus); брюшко начиная с истинного второго сегмента называется метаcома (metasoma). Стебелек исходно возник как приспособление к паразитоидному развитию личинок (см. Euhymenoptera): наличие стебелька позволяет самке манипулировать брюшком при нападении на подвижное насекомое, парализовать его уколом яйцеклада и отложить на него яйца. При этом яд, используемый для парализации, выделяется придаточной железой [см. Hexapoda (12)]. У жалящих перепончатокрылых (Monotrocha) стебелек способствует использованию жала, которое представляет собой видоизмененный яйцеклад и служит для парализации или умерщвления добычи, которой самка выкармливает личинок, а также для защиты потомства. Стебельчатое строение брюшка свойственно обоим полам, хотя используется только самкой. Стебельчатое брюшко сохраняется у всех Apocrita, в том числе и у тех, которые его не используют и утратили яйцеклад.
(2) У большинства личинок полости средней кишки (энтодермальной) и задней кишки (эктодермальной) разобщены до конца периода питания, так что выделение экскрементов происходит только перед линькой на куколку; это связано с тем, что личинки Apocrita исходно питаются жидкой гемолимфой насекомых (см. Euhymenoptera). Лишь у некоторых пчелиных кишечник стал вторично сквозным начиная с первого личиночного возраста (все пчелиные перешли к питанию пыльцой). В отличие от Apocrita, у Idiogastra сохраняется связь средней и задней кишки, несмотря на паразитоидный образ жизни.
(3) Ваннальные жилки отсутствуют; соответственно, отсутствуют поля шипиков на передних крыльях и ценхри на метанотуме [см. (6)].
2.3.3.2.1. Плезиоморфон Parasitica Hartig 1837 {synn. circ.: Pedonculiventres Latreille 1804c = Spiculifera Westwood 1840 = Entomospheces Leunis 1860 = Tristega Konov 1897; nom.hier.: Ichneumon/f1=Cynips/g6 [f:1802; g:1758(gen.213)]}. Личинки большинства видов являются паразитоидами на насекомых [см. Euhymenoptera (1)]; личинки некоторых видов паразитируют на кладках яиц насекомых, т.е. формально являются хищниками. В некоторых таксонах Parasitica личинки перешли от паразитоидного питания на насекомых-галлообразователях к самостоятельному образованию галлов на растениях и питанию тканями галла, т.е. являются настоящими фитофагами. Включает порядка 65 000 описанных видов, т.е. бóльшую часть всех видов Hymenoptera. Среди таксонов, составляющих плезиоморфон Parasitica, наиболее крупными являются Proctotrupes/f1=Cynips/g7 и Ichneumon/f2=g1; прочих Parasitica иногда объединяют в таксон Evania/fg (Rasnitsyn & Quicke 2002), либо рассматривают как несколько не связанных между собой таксонов (Grimaldi & Engel 2005).
2.3.3.2.1-1. Evania/fg [f:1802; g:1775] (incl. Stephanus, Trigonalys, Megalyra).
2.3.3.2.1-2. Proctotrupes/f1=Cynips/g7 [f:1802; g:1758(gen.213)] (incl. Eulophus). Преимущественно мелкие, с упрощенным жилкованием крыльев. Включает крупные таксоны Proctotrupes/fg [f:1802; g:1796], Cynips/fg [f:1813; g:1758], Chalcis/f=Eulophus/g [f:1817; g:1762] и др.
2.3.3.2.1-3. Ichneumon/f2=g1 [f:1802; g:L.1758(gen.215)] (incl. Bracon). Имеет непосредственное родство с Monotrocha, о чем свидетельствует синапоморфия Ichneumon/fg и Monotrocha – наличие прочного двухмыщелкового сочленения, соединяющего проподеум и метасому [см. Apocrita (1)] (Rasnitsyn & Quicke 2002, Grimaldi & Engel 2005).
2.3.3.2.2. Monotrocha Hartig 1837 (non Monotrocha Ehrenberg 1830) – жалящие {по циркумскрипции соответствует: «Aculeata» auct. (non Scopoli 1763); nom.hier.: Vespa/f7=Sphex/g1 [f:1781; g:1758(gen.216)]}. Характеризуется следующими апоморфиями. У самки имаго VIII и IX сегменты брюшка сильно модифицированы, с вестигиальными тергитами, втянуты в VII сегмент брюшка (рис. 10.7.4D); находящийся на них яйцеклад [см. Hymenoptera (15)] превращен в выдвигающееся жало и утратил функцию проведения яиц. Вертлужок [см. Hymenoptera (3)] утрачен. Антенны у самки исходно 12-члениковые, у самца исходно 13-члениковые (в некоторых таксонах на 1–2 членика меньше). Включает несколько десятков тысяч видов. В составе Monotrocha принято выделять следующие таксоны.
2.3.3.2.2-1. Плезиоморфон Scolia/fg1 [f:1802; g:1775].
2.3.3.2.2-2. Chrysis/fg1 [f:1805; g:1761].
2.3.3.2.2-3. Pompilus/fg1 [f:1805; g:1798].
2.3.3.2.2-4. Apis/f1=Sphex/g2 [f:1802; g:1758(gen.216)]. Задний край переднеспинки в медиальной части прямой, латерально резко выступает назад в виде т.н. «плечевых бугров». Включает плезиоморфон Sphex/f1=g3 [f:1802; g:1758 (gen.216)] – роющие осы и голофилетический таксон Anthophila Latreille 1807 {nom.hier: Apis/f2=g1 [f:1802, g:1758(gen.218)]} – пчелиные. Таксон Anthophila (обычно считающийся семейством или надсемейством) характеризуется наличием у самок приспособлений к сбору пыльцы цветов, которой они выкармливают своих личинок (вместо исходного выкармливания насекомыми – см. Apocrita); у некоторых видов пчелиных имеется бесплодная каста (см. ниже).
2.3.3.2.2-5. Duplicipennes Dumeril 1805 {nom.hier.: Vespa/f8=g1 [f:1781; g:1758(gen.217)] (incl. Eumenes, Masaris)} – складчатокрылые осы. У большинства представителей передние крылья в покое складываются продольно. Сюда относятся горшечные осы Eumenes/fg1 [f:1815; g:1802], цветочные осы Masaris/fg1 [f:1802; g:1793], бумажные общественные осы Vespa/f9=g2 (incl. Polistes) и некоторые другие.
2.3.3.2.2-6. Sodales Westwood 1840 {nom.typ: Formica/fg1 [f:1802; g:1758(gen.218)]} – муравьи. Второй сегмент брюшка [отделенный перетяжкой от первого – см. Apocrita (1)] отделен перетяжкой от следующего, образуя подвижный стебелек. Всегда имеется бесплодная каста, которая, в отличие от плодовитой, не имеет крыльев (см. ниже).
Выкармливание личинок и социальность среди Monotrocha. Многие представители Monotrocha перешли от паразитоидного образа жизни, исходно свойственного Euhymenoptera (см. выше) к выкармливанию личинок в специально построенном гнезде. В этом случае самка не только откладывает яйцо на жертву, но и перемещает эту жертву из ее естественного места обитания в гнездо, приспособленное для развития личинки. В наиболее примитивном случае самка лишь перемещает парализованную добычу в естественное укрытие и откладывает на нее яйцо, так что далее личинка развивается как эктопаразитоид. Во многих случаях самка сначала делает гнездо определенной видоспецифичной конструкции, затем разыскивает добычу, парализует ее, перемещает в гнездо, откладывает на жертву яйцо и запечатывает гнездо. У некоторых видов самка помещает в гнездо не одно, а несколько парализованных насекомых; таким образом, здесь, в отличие от исходного паразитоидного питания, возможно питание личинок более мелкими жертвами. У некоторых видов самка не запечатывает гнездо, а на протяжении всего развития личинок выкармливает их свежеубитыми насекомыми или иной пищей животного происхождения. В отдельных случаях произошел переход от выкармливания личинок животной пищей к выкармливанию иной пищей: в частности пчелы (Anthophila, или Apis/fg) и цветочные осы (Masaris/fg) выкармливают личинок не насекомыми, а пыльцой растений; большинство муравьев (Sodales, или Formica/fg) выкармливают личинок насекомыми, но некоторые муравьи перешли к выкармливанию личинок семенами растений, а муравьи-листорезы (Atta/fg) кормят личинок специально разводимыми грибами. Во всех случаях все функции, связанные заботой о потомстве (постройка гнезда, добывание пищи, защита незапечатанного гнезда и др.) выполняются только самкой; несмотря на то, что в некоторых случаях эти работы бывают трудоемкими, самцы никогда не принимают в них участия; то есть настоящих семей у перепончатокрылых не бывает (в отличие от некоторых других насекомых со сложным поведением, у которых возникают постоянные семейные пары). Вероятно, это вызвано тем, что исходно основные родительские функции связаны с наличием жалящего яйцеклада, который используется и как средство добывания пищи для личинок, и как средство их защиты, а яйцеклад может быть развит только у самок.
Вместо этого среди Monotrocha, выкармливающих личинок в гнезде, имеется большое число социальных видов. Одна и та же особая форма социальности, присущая Monotrocha, встречается у представителей разных таксонов, то есть возникала в ходе эволюции независимо несколько раз. При этой форме социальности все самки колонии разделены на плодовитую и бесплодную касты; в отличие от других социальных животных (термиты и др.), у социальных перепончатокрылых на касты делятся только самки, тогда как все самцы являются обычными плодовитыми особями и не участвуют в функционировании колонии. Многократное возникновение такой социальности вызвано сочетанием двух причин: 1) для выполнения сложных функций по выращиванию потомства (добыванию пищи, защиты и др.) самкам выгодно кооперироваться друг с другом, поскольку самцы в этом участвовать не могут; 2) как и все перепончатокрылые, эти насекомые имеют гапло-диплоидное определение пола, и поэтому самка имеет бóльшее генетическое сходство со своими сестрами, чем с матерью [см. Hymenoptera (10)]. Это приводит к тому, что естественный отбор может быть направлен на поддержание такого поведения, которое способствует выращиванию своих сестер в ущерб выращиванию своих дочерей.
* * *