КЛАДОЭНДЕЗИС НАСЕКОМЫХ

![]()
|
КЛАДОЭНДЕЗИС НАСЕКОМЫХ |
|||||||
|
|
|
fg | |||||
X-1.3.4.2. Lepidoptera Linnaeus 1758 – чешуекрылые, или бабочки
(Somobialia![]() Eucaryota
Eucaryota![]() Metazoa
Metazoa![]() Monostomata
Monostomata![]() Haemataria
Haemataria![]() Peritoneata
Peritoneata![]() Metameria
Metameria![]()
Gnathopoda![]() Euarthropoda
Euarthropoda![]() Mandibulata
Mandibulata![]() Atelocerata
Atelocerata![]() Hexapoda
Hexapoda![]() Amyocerata
Amyocerata![]()
Pterygota![]() Metapterygota
Metapterygota![]() Neoptera
Neoptera![]() Eumetabola
Eumetabola![]() Metabola
Metabola![]() Mecopteriformia
Mecopteriformia![]() Amphiesmenoptera
Amphiesmenoptera![]() Lepidoptera)
Lepidoptera)
|
Названия, однозначно соответствующие по циркумскрипции: — Lepidoptera Linnaeus 1758; — Hymenolepidoptera Schaeffer 1766; — Farinacea = Farinosa Schluga 1767; — Glossata Fabricius 1775; — Roeselii Scopoli 1777; — Lepidioptera Clairville 1798; — Glossostoma Leach 1815a; — Lepidopteria Rafinesque 1815; — Glossota Amyot 1845; — Micrognatha Spinola 1850; — Sorbentia Haeckel 1889; — Lepidopterida Pearce 1936. По циркумскрипции также соответствует таксонам под типифицированными названиями, образованными от родового названия Papilio: Papilionoides, Papilionida. Иерархическое название: Papilio/fg3 [f:1781; g:L.1758(gen.203)] (sine Phryganea; incl. Micropterix). |
|
Ранг. Обычно отряд.
Название. В публикации, считающейся исходной (Linnaeus 1758), в состав отряда Lepidoptera, помимо прочих видов, был включен вид Phalaena Tinea proletella, ныне относимый не к Lepidoptera, а к Scytinelytra в составе Arthroidignatha. Этот вид был включен Линнеем в Lepidoptera явно по ошибке, поскольку его комплексный ротовой аппарат был принят за простой хоботок бабочек («lingua inflexa»), а восковой налет на крыльях – за чешуйки. Таким образом, название Lepidoptera следует считать соответствующим по исходной циркумскрипции данному таксону, не включающему proletella [Phalaena] (см. I.6.5.1).
Аутапоморфии Lepidoptera
(1) Чешуйки. Щетинки, покрывающие крылья [см. Amphiesmenoptera (4)], преобразованы в чешуйки; у имаго все тело, включая крылья, покрыто сплошным покровом из черепицеобразно налегающих друг на друга чешуек [представляющих собой уплощенные и расширенные щетинки – см. Euarthropoda (5c)]. Благодаря чешуйчатому покрову крылья выглядят непрозрачными и интенсивно окрашенными (с чем связано название Lepidoptera – чешуекрылые); у многих бабочек разноцветные чешуйки образуют видоспецифичные узоры. Сплошной чешуйчатый покров имеется у подавляющего большинства бабочек. У некоторых видов на крыльях имеются более или менее крупные прозрачные участки, лишенные чешуек и волосков, так что крылья кажутся голыми, но в таких случаях сплошной чешуйчатый покров сохраняется в виде каймы по краю крыла и вдоль жилок. В редких случаях чешуйки сужены и закрывают не всю поверхность крыла, так что крыло выглядит полупрозрачным.
Каждая чешуйка имеет узкое основание, которым она подвижно крепится в своем гнезде; на большем своем протяжении чешуйка пластинчато расширена, и ее поверхность исчерчена многочисленными параллельными продольными ребрышками.
Как и всякое производное щетинки, чешуйка бабочки состоит из кутикулы, выделенной оболочечной трихогенной клеткой, исходно окружающей сенсорный нейрон; но в отличие от щетинок и чешуек других насекомых, чешуйки бабочек утратили иннервацию. По крайней мере для одного из видов бабочек показано, что при развитии чешуйки (которое происходит под покровом куколочной кутикулы) гиподермальная клетка-предшественник делится на крупную дистальную клетку (которая в дальнейшем делится на трихоген и тормоген) и маленькую проксимальную клетку, которая в дальнейшем отмирает (Stossberg 1938); по-видимому, эта отмирающая клетка соответствует общему предшественнику нейрона и текогена [см. Euarthropoda (5b)] (Keil 1997).
Чешуйки, сходные с чешуйками бабочек, имеются у многих других насекомых [см. Euarthropoda (5c)].
Высказывались предположения, что чешуйчатый покров на крыльях бабочек влияет на потоки воздуха у поверхности крыла и тем самым способствует улучшению полета. Однако это не позволяет объяснить факт наличия чешуек у всех бабочек, имеющих разнообразное строение крыльев и разные способы полета.
У наиболее примитивных бабочек крылья по форме и жилкованию сходны с крыльями примитивных ручейников и сохраняют черты их общего предка [см. Amphiesmenoptera (5)]. Исходно бабочки бимоторные, с почти одинаковыми передними и задними крыльями (рис. 10.16.1); в полете крылья не сцеплены, так что передние и задние крылья работают самостоятельно. Такие гомономные крылья сохраняются у Palaeolepidoptera, Protolepidoptera и многих Neolepidoptera-Exoporia. У Frenatae, куда относится подавляющее большинство видов, крылья исходно переднемоторные (см. ниже, Neolepidoptera: «Классификация»: Frenatae). Характер полета у разных бабочек очень различен. Дневные бабочки (Rhopalocera), имеют особый порхающий тип полета: их широкие крылья последовательно схлопываются над и под телом, каждый раз выбивая струю воздуха назад. У бражников (Sphinx/fg), имеющих узкие крылья и крупную среднегрудь с мощными крыловыми мышцами, полет похож на полет перепончатокрылых: с большой частотой и малой амплитудой взмахов. Между этими крайними типами полета имеется множество других. Наиболее примитивные бабочки имеют, как правило, молевидный облик: мелкие, с небольшими узкими крыльями и слабым полетом. Таковы большинство Protolepidoptera, Palaeolepidoptera, Exoporia-Mnesarchaea/fg, Frenatae-Aculeatae, а из Frenatae-Ditrysia – настоящие моли (Tinea/fg) и многие другие. Отсюда можно сделать вывод, что чешуйки возникли у общего предка Lepidoptera на фоне мелких размеров и слабого полета, а сохранились у всех видов этого гигантского таксона на фоне крайнего разнообразия формы и функции крыльев. У подавляющего большинства бабочек крылья развиты и используются для полета, лишь у самок немногих видов крылья вестигиальные или утрачены; в этих случаях чешуйки также сохраняются.
У большинства бабочек чешуйки имеют сложное строение: верхняя и нижняя пластинки, составляющие чешуйку, разделены, так что между ними находятся более или менее выраженные полости; верхняя пластинка (обращенная от поверхности крыла) имеет отверстия, соединяющие эти полости с внешней средой; эти отверстия располагаются между продольными ребрами; апикальный край такой чешуйки зазубренный, т.е. с выемками между вершинами ребер. Такое строение чешуек свойственно Neolepidoptera, Archifrenata, Acanthopteroctetes/fg и Lophocorona/fg. Наряду с такими чешуйками на крыле могут присутствовать и чешуйки более простого строения – без полости, без отверстий и с ровными краями (Simonsen 2001). Некоторые авторы считают наличие чешуек с полостями не исходным для Lepidoptera, а аутапоморфией таксона Coelolepida (см. ниже, Palaeolepidoptera: «Статус и классификация»).
(2) Утрата медиального глазка. У имаго медиальный глазок всегда утрачен, так что бывают развиты лишь 2 глазка, либо простые глазки полностью утрачены. То же у многих других насекомых, в том числе у некоторых Trichoptera [см. Amyocerata (4)]. Фасеточные глаза обычно хорошо развиты; лишь у некоторых бескрылых безногих самок глаза вестигиальные или утрачены.
(3) Модификация максиллярных щупиков. У имаго максиллярный щупик [5-члениковый – см. Hexapoda (2c)] имеет характерную форму: сочленение 2-го и 3-го члеников неподвижное, 4-й членик удлинен, 5-й членик с вогнутостью, в которой находятся сенсиллы; щупик имеет два особенно подвижных сочленения – между 1-м и 2-м и между 3-м и 4-м члениками, так что обычно сгибается Z-образно (рис. 10.16.1). Примитивные бабочки, имеющие грызущий ротовой аппарат и питающиеся пыльцой [см. ниже, плезиоморфон Protolepidoptera (1)] используют максиллярные щупики такого строения для собирания пыльцы (Scoble 1992). Помимо Protolepidoptera, такое же строение щупика сохраняется и у некоторых Glossolepidoptera – у Palaeolepidoptera и немногих Neolepidoptera, несмотря на то, что они утратили способность питаться пыльцой. У большинства Neolepidoptera максиллярный щупик имаго в той или иной степени редуцирован, с уменьшенным числом члеников, или совсем утрачен.
(4) Эпифиз. На передней голени срединная шпора [см. Amphiesmenoptera (7)] преобразована в придаток, называемый эпифиз (epiphysis) и служащий для чистки антенн: она расширена и резко отличается от других шпор; апикальные шпоры передней голени утрачены (рис. 10.16.1). У отдельных представителей эпифиз утрачен.
Признаки Lepidoptera неясного филогенетического значения
(5) Форма мандибул куколки. У наиболее примитивных чешуекрылых куколки несклеенные, с действующими мандибулами [«pupa dectica» – см. Amphiesmenoptera (10)]. При этом как среди чешуекрылых с грызущим ротовым аппаратом имаго (Protolepidoptera), так и среди хоботных чешуекрылых (Glossolepidoptera) есть представители, куколки которых имеют необычное строение мандибул, которое можно назвать «лепидоптероидным». Такая лепидоптероидная мандибула очень сильно удлинена, в проксимальной части имеет форму изогнутого стержня, к вершине раструбообразно расширяется и заканчивается вогнутой округлой площадкой, обрамленной зубцами. Помимо этих вершинных зубцов бывает развит еще один крупный зубец на наружной стороне мандибулы приблизительно в 1/3 от ее вершины (у Aglossata такой предвершинный зубец имеется на обеих мандибулах, у Heterobathmia/fg – только на правой мандибуле, у Palaeolepidoptera предвершинные зубцы отсутствуют). В сложенном состоянии такие мандибулы перекрещиваются своими изогнутыми основаниями, а их расширенные зазубренные вершины направлены в противоположные стороны и выступают за латеральные края головы. Из-за своего огромного размера лепитоптероидные мандибулы непригодны для кусания; вероятно, они исходно специализированы для рытья. Так, куколка Eriocrania/fg cicatricella [Adela] (= purpurella [Tinea]) использует мандибулы для того, чтобы выбраться из-под земли (где происходило ее развитие) на поверхность почвы, где она линяет на имаго: энергично раздвигая и сдвигая мандибулы и поднимая и опуская голову, куколка (точнее, фаратное имаго) перемещается в толще почвы, не двигая ногами (Chapman 1893). Среди Protolepidoptera лепидоптероидные куколочные мандибулы имеются у Aglossata и Heterobathmia/fg (Dumbleton 1952, Kristensen & Nielsen 1982); в отличие от них у Micropterix/fg (incl. Sabatinca) куколочные мандибулы не трубообразные, имеют обычный размер и треугольно-заостренную форму (Tillyard 1923, Hashimoto 2006). Среди Glossolepidoptera лепидоптероидные мандибулы имеются у всех известных куколок Palaeolepidoptera; у всех Neolepidoptera (куда относится подавляющее большинство чешуекрылых) куколочные мандибулы редуцированы [см. ниже, Neolepidoptera (1)].
Уникальная лепидоптероидная форма куколочных мандибул, несомненно возникла в ходе эволюции однократно. Относительно того, аутапоморфией какого именно таксона является этот признак, возможны следующие два предположения. (1) Лепидоптероидная форма мандибул – это аутапоморфия Lepidoptera; в этом случае у Micropterix/fg (incl. Sabatinca) куколочные мандибулы вторично упростились, став похожими на мандибулы куколок ручейников и других «pupa dectica». (2) Лепидоптероидная форма мандибул – это аутапоморфия таксона, объединяющего Aglossata, Heterobathmia/fg и Glossolepidoptera; в этом случае у Micropterix/fg (incl. Sabatinca) куколочные мандибулы сохраняют строение, исходное для Amphiesmenoptera (см. ниже, Protolepidoptera «Статус и классификация»).

(6) Растительноядные личинки. Личинки подавляющего большинства видов фитофаги и адаптированы к жизни на растениях. Личинки всех Lepidoptera сохраняют грызущий ротовой аппарат с развитыми мандибулами [см. Amphiesmenoptera (12)]. Многие из них живут открыто на растениях и имеют развитые, но обычно очень короткие грудные ноги с полным набором члеников и брюшные ложноножки [см. ниже, Neolepidoptera (2)]. Некоторые личинки перешли к скрытому образу жизни, в этом случае ноги бывают вестигиальными или утрачены, а ложноножки также отсутствуют (это личинки всех известных Protolepidoptera, Palaeolepidoptera и некоторых Neolepidoptera).
Подавляющее большинство личинок чешуекрылых обитает в воздушной среде. Лишь у немногих видов Neolepidoptera личинки перешли к жизни в воде: их трахеостии замкнуты (но в последнем личиночном возрасте могут становиться дыхальцами); некоторые водные личинки имеют нитевидные трахейные жабры. В отличие от личинок ручейников и большинства других гидробионтов, водные личики чешуекрылых являются настоящими фитофагами и питаются живыми частями водных сосудистых растений.
Помимо фитофагов, среди личинок бабочек встречаются детритофаги, микофаги, хищники, паразитоиды, и даже кератофаги [настоящие моли – Tinea/fg (incl. Monopis)]. Питание насекомыми (хищничество и паразитоидность) для гусениц бабочек явно вторично; оно возникало неоднократно, но общее число хищных гусениц ничтожно мало по сравнению с общим числом видов. По мнению некоторых авторов детритофагия и микофагия могут быть исходными для личинок бабочек, поскольку они исходны для насекомых в целом. Однако наличие настоящих фитофагов во всех главных филогенетических ветвях Lepidoptera свидетельствует в пользу того, что фитофагия личинок была приобретена общим предком бабочек и является исходной для всего этого таксона.
Геологический возраст. Юра – ныне (Rasnitsyn & Quicke 2002). По жилкованию крыльев Lepidoptera не отличаются от других Amphiesmenoptera, поэтому некоторые авторы указывают более древней возраст Lepidoptera, вплоть до пермского (см. Amphiesmenoptera).
Классификации Lepidoptera
I. Деление Lepidoptera на Heterocera и Rhopalocera
Таксон Rhopalocera Dumeril 1805 {syn.obj.: Globulicornes Dumeril 1805; synn.circ.: Diurna Latreille 1809 (non Diurni Latreille 1802b) = Ropaloceria Rafinesque 1815 = Ephemerina Billberg 1820 = Clavicornes Latreille 1825 (non Clavicornes Dumeril 1800) = Eulepidoptera Crampton 1916 = Rhopalocerina Pearce 1936 = Aparasternaria Niculescu 1970; по циркумскрипции также соответствует роду Papilio sensu Linnaeus 1758; nom.typ.: Papilio/fg (incl. Hespria)} – булавоусые, или дневные бабочки. У имаго антенны булавовидные, т.е. утолщаются к вершине; крылья утратили способность складываться на спине [см. Neoptera (1)], так что в покое либо поняты вверх и обращены вентральной поверхностью латерально, либо распластаны в стороны; в отличие от большинства других бабочек, Rhopalocera ведут дневной образ жизни и не прилетают ночью на свет. По мнению многих современных авторов этот таксон является голофилетическим.
Таксон Heterocera Boisduval 1836 {syn.circ.: Parasternaria Niculescu 1970} объединяет всех остальных чешуекрылых, характеризуется только плезиоморфиями и является парафилетическим; по числу видов он значительно превосходит таксон Rhopalocera. В настоящее время таксон Heterocera не признают, а таксон Rhopalocera рассматривают не как прямо подчиненный Lepidoptera, а в составе более дробного таксона внутри Lepidoptera.
II. Деление Lepidoptera на Microlepidoptera и Macrolepidoptera
Таксон Macrolepidoptera auct. {таксоны близкие по циркумскрипции: Macrolepidoptera sensu Gerstaecker 1863, Macrofrenatae Comstock 1893, Eulepidoptera Crampton 1916a, Macroptera Börner 1920} объединяет дневных и крупных ночных бабочек. Авторство названия «Macrolepidoptera» не установлено. В настоящее время некоторые авторы используют название «Macrolepidoptera» в значении, близком к традиционному, для голофилетического таксона в составе Ditrysia (см. ниже, Neolepidoptera: «Классификация»).
Таксон Microlepidoptera auct. объединяет всех остальных чешуекрылых и является парафилетическим.
III. Деление Lepidoptera на Monotrysia и Ditrysia
Таксон Ditrysia Börner 1925 характеризуется тем, что самка имеет два непарных генитальных отверстия (яйцекладное и копулятивное), соединенных внутренним протоком. Ditrysia представляет собой голофилетический таксон, включающий подавляющее большинство чешуекрылых (см. ниже, Neolepidoptera: «Классификация»).
Таксон Monotrysia Börner 1925 характеризуется иным строением гениталий самки: либо имеются два отверстия, но без внутреннего протока (у Exoporia – см. ниже, Neolepidoptera: «Классификация»), либо имеется единственное половое отверстие, служащее и для копуляции, и для яйцекладки (как у большинства других насекомых). Таксон Monotrysia является парафилетическим и поэтому в современных классификациях не признан.
IV. Деление Lepidoptera на Frenatae и Jugatae
Таксон Frenatae Comstock 1892 характеризуется апоморфией – редукцией югума и наличием особой френатной сцепки, обеспечивающей механическое соединение переднего и заднего крыла в полете; в современных классификациях Frenatae рассматривают как подчиненный таксон в составе Neolepidoptera (см. ниже, Neolepidoptera: «Классификация»).
Таксон Jugatae Comstock 1892 {synn.circ.: Isoneuria Karsch 1898 = Archilepidoptera Spuler 1910 = Homoneura Tillyard 1917d (non Homoneura Enderlein 1903); nom.typ.: Hepialus/fg (incl. Micropterix)}. Характеризуется отсутствием френатной сцепки заднего крыла с передним и наличием развитого югума. Апоморфией этого таксона считалось наличие особой югатной сцепки; однако у многих представителей крылья вообще не способны сцепляться, а наличие югума является плезиоморфией [см.Amphiesmenoptera (4) и Metabola (4)]. В настоящее время эту группу считают парафилетической и не признают в качестве таксона.
V. Деление Lepidoptera на Frenatae, Jugo-frenata и Macrojugatae. В первой половине XX века было широко принято делить таксон Jugatae на Microjugatae и Macrojugatae; иногда Microjugatae и Macrojugatae рассматриваются как таксоны того же ранга, что и Frenatae.
Таксон Frenatae (см. IV и ниже, Neolepidoptera: «Классификация»).
Таксон Jugo-frenata Tillyard 1918b {= Microjugatae s.l.; nom.typ.: Micropterix/fg (incl. Eriocrania)}. Особенностью этого таксона считалось наличие юго-френатной сцепки, при которой югум переднего крыла подогнут вентрально и сцепляется с щетинками, представляющими собой френулюм заднего крыла [см. Metabola (4)] (Tillyard 1918b–1919a). Однако в действительности югум представляет собой инверталяре [см. Neoptera (1a)], так что при расправленных крыльях югум переднего крыла не подогнут, а расправлен и не обеспечивает сцепку (рис. 10.16.1). В настоящее время таксон Jugo-frenata не признан, поскольку является парафилетическим; вместо него рассматривают отдельно Protolepidoptera (см. ниже), Palaeolepidoptera (см. ниже) и Mnesarchaea/fg (см. ниже, Neolepidoptera: «Классификация»).
Таксон Macrojugatae Comstock 1893 {nom.typ.: Hepialus/fg (incl. Prototheora)}. Особенностью этого таксона считалось наличие настоящей югатной сцепки, при которой передний край заднего крыла зажат между расправленным югумом, прижатым к нему с вентральной стороны, и продолжением заднего края переднего крыла, прижатым к нему с дорсальной стороны (Tillyard 1918b–1919a). Однако у многих Macrojugatae югум мягкий и не обеспечивает сцепку. В современных классификациях Macrojugatae рассматривается как подчиненный таксон в составе Neolepidoptera (см. ниже).
VI. Деление Lepidoptera на Protolepidoptera и Glossolepidoptera. В этой классификации таксон Protolepidoptera характеризуется плезиоморфией – грызущим ротовым аппаратом у имаго (см. ниже), а таксон Glossolepidoptera – специализированным сосущим ротовым аппаратом особого строения, что свидетельствует о его голофилии (см. ниже). Помимо типичных Protolepidoptera и типичных Glossolepidoptera, существуют две малочисленные группы бабочек, занимающих спорное систематическое положение – это Aglossata и Heterobathmia/fg; иногда их рассматривают как таксоны того же ранга, что Protolepidoptera и Glossolepidoptera (Kristensen 1984); иногда включают в Protolepidoptera s.l. на основании симплезиоморфий (см. ниже, Protolepidoptera: «Статус и классификация»).
В упрощенном виде филогения Lepidoptera и соотношение классификаций I–VI выглядят следующим образом:
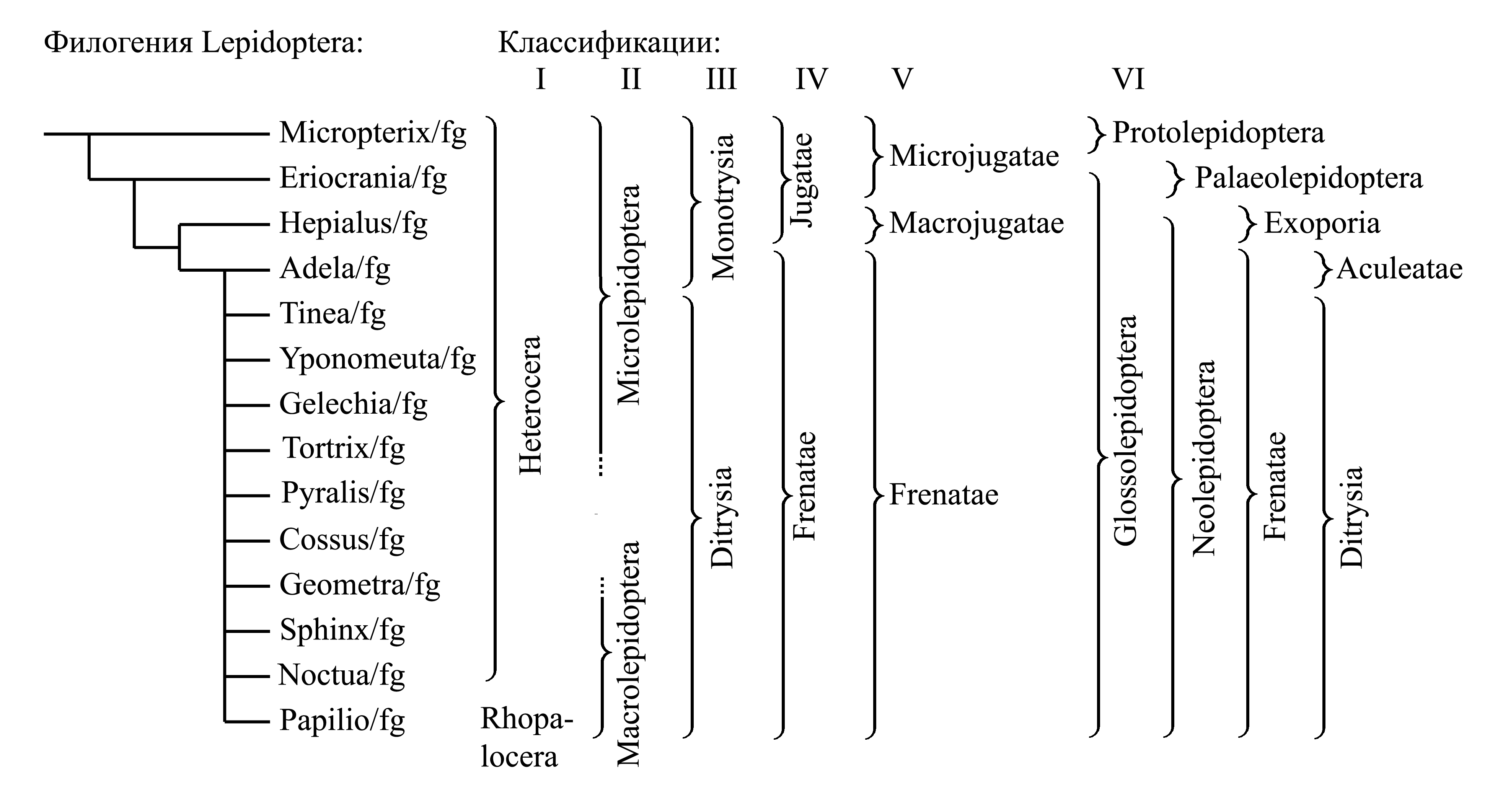
Здесь мы принимаем деление Lepidoptera на плезиоморфон Protolepidoptera и голофилетический таксон Glossolepidoptera.
Lepidoptera, или Papilio/fg3
![]() 1. Плезиоморфон Protolepidoptera,
или Micropterix/fg1
1. Плезиоморфон Protolepidoptera,
или Micropterix/fg1
![]() 2. Glossolepidoptera,
или Papilio/fg4
2. Glossolepidoptera,
или Papilio/fg4
![]() 2.1. Плезиоморфон Palaeolepidoptera,
или Eriocrania/fg1
2.1. Плезиоморфон Palaeolepidoptera,
или Eriocrania/fg1
![]() 2.2. Neolepidoptera, или
Papilio/fg5
2.2. Neolepidoptera, или
Papilio/fg5