КЛАДОЭНДЕЗИС НАСЕКОМЫХ

![]()
|
КЛАДОЭНДЕЗИС НАСЕКОМЫХ |
|||||||
|
|
|
fg | |||||
X-1.3.4.1. Trichoptera Kirby 1813 – ручейники
(Somobialia![]() Eucaryota
Eucaryota![]() Metazoa
Metazoa![]() Monostomata
Monostomata![]() Haemataria
Haemataria![]() Peritoneata
Peritoneata![]() Metameria
Metameria![]()
Gnathopoda![]() Euarthropoda
Euarthropoda![]() Mandibulata
Mandibulata![]() Atelocerata
Atelocerata![]() Hexapoda
Hexapoda![]() Amyocerata
Amyocerata![]()
Pterygota![]() Metapterygota
Metapterygota![]() Neoptera
Neoptera![]() Eumetabola
Eumetabola![]() Metabola
Metabola![]() Mecopteriformia
Mecopteriformia![]() Amphiesmenoptera
Amphiesmenoptera![]() Trichoptera)
Trichoptera)
(рис. 10.15.1–10.15.4)
(рис. 10.15.1, 10.15.2A–C,
10.15.2D–F, 10.15.3,
10.15.4)
|
Названия, однозначно соответствующие по циркумскрипции: — Trichoptera Kirby 1813; — Gnathostoma Leach 1815a; — Plicipennes Latreille 1817; — Lophiacera Billberg 1820; — Trichopterida Pearce 1936. По циркумскрипции также соответствует таксонам под типифицированными названиями, образованными от родового названия Phryganea: Phryganites, Phryganida, Phryganidia, Phryganeoides, Phryganina, Phryganidae, Phryganeodea, Phryganinae, Phryganeides, Phryganodea, Phryganeidae, Phryganaria, Phryganoidea, Phrydaneida. Иерархическое название: Phryganea/fg1 [f:1804; g:L.1758(gen. 209)] (incl. Philopotamus). |
|
Ранг. В XX веке – обычно отряд; в первой половине XIX века – от рода до отряда.
Аутапоморфии Trichoptera
(1) Амфибиотизм и особенности куколки. Личинки и куколки перешли к обитанию под водой, тогда как имаго сохранили наземный образ жизни; при этом у куколки появилось уникальное приспособление к выходу из воды: куколочные ноги всех трех пар по-разному модифицированы [см. (1a)], благодаря чему в состоянии фаратного имаго куколка способна активно плавать и выбираться на сушу [см. (1b)]. В связи с подводным образом жизни личинок и куколок их трахейная система адаптирована к дыханию растворенным в воде кислородом [см. (1c)]. У подавляющего большинства видов ручейников личинки и куколки обитают в пресных водах; у Австралийско-Новозеландского вида Philanisus plebeius Walker 1852 личинки и куколки перешли к жизни в морской среде; у немногих видов ручейников личинки и куколки вторично приспособились к жизни на суше и обитают во влажной лесной подстилке.
(1 a) Модификации в строении куколки. Ноги куколки дифференцированы следующим образом: на передней и средней ноге претарсус несет два неподвижных склеротизованных крючка, соответствующих имагинальным коготкам (рис. 10.15.4C); на средней ноге лапка несет длинные густые щетинки (рис. 10.15.4D); на задней ноге таких таких щетинок нет, а крючки претарсуса отсутствуют или развиты хуже, чем на передней и средней ногах (рис. 10.15.4E). Благодаря этому передние и средние ноги куколки могут использоваться как ходильные (с цепкими коготками), а средние – также как плавательные (с плавательными щетинками); задние ноги непригодны ни для того, ни для другого. Ноги куколки могут функционировать только в состоянии фаратного имаго, поскольку до этого ножные мышцы отсутствуют, и все ноги неподвижно согнуты в коленях [см. Metabola (1Ccδ) и (1Ccδ-11)] (рис. 10.15.3С).
(1 b) Развитие и поведение куколки. Обитающая под водой личинка последнего возраста [см. Metabola (1A)], закончив питание, строит убежище для последующего превращения в куколку – куколочный домик; этот домик неподвижно прикреплен к подводному предмету и обычно состоит из тех или иных твердых частиц (песчинок, фрагментов растительных остатков или других), скрепленных шелком, который личинка выделяет лабиальными железами [см. Mecopteriformia (3)]. Личинки Integripalpia, постоянно живущие в переносном личиночном домике, перестраивают свой личиночный домик в куколочный: прикрепляют его к субстрату и заделывают выходное отверстие. У Spicipalpia личинка внутри домика окружает себя непроницаемым эластичным шелковым коконом, внутри которого образуется осмотическое давление; газообмен, необходимый для ее дыхания, происходит только сквозь стенки этого кокона. Возможно, наличие такого кокона является исходным для ручейников (Wiggins & Wichard 1989). У Curvipalpia и Integripalpia личинка не делает сплошного кокона, так что свободно омывается водой, которая проходит через переднее и заднее отверстия домика; в этом случае личинка, а в последующем куколка, способна создавать ток воды спереди назад с помощью волнообразных движений брюшка.
Внутри этого убежища личинка впадает в состояние пронимфы и затем линяет на куколку; происходящая при этом трансформация ног приводит к тому, что ноги куколки полностью утрачивают мышцы [см. Metabola (1Ccδ-11) и рис. 10.15.2A–C, 10.15.2D–F, 10.15.3]. У куколки, находящейся в домике, ноги неподвижны и имеют обычную позу: плотно согнуты в коленях и повернуты так, что колени передних и средних ног выступают впереди протоптеронов, а колени задних ног скрыты под протоптеронами (рис. 10.15.3С) [см. Metabola (1Ba)].
У большинства ручейников куколка имеет крупные подвижные склеротизованные куколочные мандибулы [в отличие от вестигиальных мандибул имаго – см. (4)], т.е. куколка относится к типу «pupa dectica» [точнее, «pupa neodectica» – см. Amphiesmenoptera (10) и Metabola (1Bb)]; перед линькой на имаго такая куколка прогрызает мандибулами кокон или переднюю стенку домика. Лишь у некоторых видов мандибулы куколки вторично редуцированы: уменьшены, не склеротизованы и не способны функционировать (т.е. куколка относится к типу «pupa adectica»); в этих случаях личинка строит куколочный домик без прочной передней стенки (Wiggins 1960).
После того, как под куколочной кутикулой сформируются имагинальная кутикула и имагинальные мышцы (т.е. в состоянии фаратного имаго) куколка приобретает способность активно плавать с помощью средних ног [снабженных щетинками – см. (1a)] и ползать с помощью передних и средних ног [снабженных коготками – см. (1a)] – т.е. куколка относится к типу «pupa neogressibilis» [см. Metabola (1Bb)]. При этом задние ноги, колени которых закрыты протоптеронами, остаются неподвижными. Выбравшись из кокона или домика, куколка плывет под водой, будучи ориентированной вентральной стороной вверх и делая синхронные гребные движения средними ногами (рис. 10.15.4A). Достигнув берега или выступающего из воды предмета, куколка залезает на него при помощи передних и средних ног и, оказавшись в воздушной среде, линяет на имаго.
Таким образом, фаратное имаго (т.е. насекомое, имеющее одновременно и куколочную, и имагинальную кутикулы) по своему поведению резко отличается и от собственно куколки (которая не двигает ногами), и от имаго (которое не двигает мандибулами и не использует средние ноги для подводного плавания).
(1 c) Дыхательная система личинок и куколок. У личинок всех возрастов и у куколки трахейная система замкнутая (апнейстическая), т.е. все трахеостии [10 пар – см. Amyocerata (2)] не функционируют в качестве дыхалец и снаружи могут быть не видны. У подавляющего большинства ручейников личинки обитают в воде; лишь у немногих видов личинки вторично перешли к жизни на суше, но их трахейная система остается замкнутой. Трахейная система замкнута так же у многих других водных насекомых (см. Указатель признаков [4.2.4]), но в данном случае этот признак является аутапоморфией, поскольку в сестринском таксоне Lepidoptera трахейная система личинок стала замкнутой лишь у немногих представителей и то, как правило, не во всех возрастах.
Личинки и/или куколки многих видов ручейников имеют трахейные жабры, которые представляют собой простые или ветвистые мягкие тонкостенные выросты на тех или иных частях тела; они могут начинаться парой стволов на каждом сегменте (рис. 10.15.1C), либо порознь отходят от поверхности тела. В обоих случаях эти трахейные жабры могут быть придыхальцевыми, снабжаясь трахеями, идущими от пары дыхальцевых трахей того же сегмента тела. Вероятно, трахейные жабры неоднократно возникали в разных таксонах ручейников, как и придыхальцевые трахейные жабры многих других насекомых [см. Hexapoda (9) и Metapterygota (6)].
Вероятно, с водным образом жизни личинок связано преобразование их пигопода в пару кручьев [см. (2)].
(2) Модификация пигопода и локомоторный аппарат личинок. У личинки на десятом сегменте брюшка [последнем – см. Hexapoda (8)] прикрепительный орган [пигопод – см. Metabola (1Abα)] представляет собой пару склеротизованных крюков, направленных назад и загнутых вентрально; каждый крюк приводится в движение антагонистическими мышцами – сгибателями и разгибателями, крепящимися внутри того же десятого сегмента брюшка (рис. 10.15.1C–D). У личинок Integripalpia, обитающих в переносных домиках, десятый сегмент брюшка округлый, и крюки сидят на его поверхности по бокам от анального отверстия; при передвижении личинка выдвигает из домика голову и грудь с ногами, а крюки использует для удержания домика на брюшке. У свободноживущих личинок Isopalpoidea десятый сегмент брюшка раздвоен и образует пару вытянутых назад отростков – анальных ложноножек, между которыми располагается анальное отверстие; в этом случае крюки сидят на вершинах этих ложноножек (рис. 10.15.1C–D); при передвижении такая личинка пользуется ногами и анальными ложноножками. Анальная ложноножка может иметь склериты, придающие ей сходство с членистой ногой. Некоторые авторы интерпретируют анальные ложноножки личинок ручейников как гомологи ног с коготком на вершине; такая гомологизация неправдоподобна, поскольку у всех насекомых абдоминальные придатки не имеют коготков [см. Hexapoda (7)], а коготки ног не имеют мышц-разгибателей [см. Atelocerata (3)]. Помимо ручейников, парные крюки пигопода имеются у водных личинок некоторых других насекомых с полным превращением (см. Указатель признаков [3.47]).
В отличие от личинок Lepidoptera и некоторых других насекомых с полным превращением, у личинок ручейников I–IX сегменты брюшка не имеют ложноножек [см. Metabola (1Abβ)]. Ноги всех трех пар хорошо развиты, с полным набором члеников [см. Amphiesmenoptera (13)] и одним коготком [см. Mecopteriformia (4)]; в отличие от Lepidoptera, ноги сравнительно длинные.
(3) Редукция антенн личинки. Личиночная антенна крайне уменьшена, нерасчлененная; утрачены не только мышцы скапуса [см. Metabola (1Cb)], но и мышцы головы, идущие к основанию антенны (в отличие от личинок Lepidoptera, у которых антенна может иметь до трех члеников и двигается мышцами, подходящими к ее основанию).
(4) Модификация ротового аппарата имаго. У имаго ротовой аппарат лижуще-сосущий (рис. 10.15.1E): мандибулы утрачены или сохраняются в виде нефункционирующих вестигиев [в отличие от куколочных – см. (1b)]; функционирующей частью ротового аппарата является короткий хоботок, или гаустеллум (haustellum), образованный гипофаринксом; вершина гаустеллума мембранозная, способна вворачиваться (Crichton 1957). Такой ротовой аппарат адаптирован для слизывания жидкой пищи; имаго ручейников питаются либо нектаром цветков и/или сахаристыми выделениями членистохоботных, либо потребляют только воду, либо вообще не питаются.
Максиллы, максиллярные щупики и лабиальные щупики либо сохраняют плезиоморфные черты, либо модифицированы по-разному у разных ручейников. Щупики обычно развиты: максиллярные щупики могут сохранять все 5 члеников, а лабиальные – все 3 членика (рис. 10.15.1E) [см. Hexapoda (2) и (3)]; у самцов максиллярные щупики бывают специализированы, с уменьшенным числом члеников; иногда максиллярные и лабиальные щупики вестигиальные или утрачены. У Dipseudopsis/fg дистальные доли максилл (соответствующие лациниям или галеам) сильно удлинены, приспособлены для сосания нектара, и напоминают хоботок бабочек [ср.: Glossolepidoptera (1)].
По общему направлению специализации имагинального ротового аппарата (редукция мандибул и приспособление к сосанию нектара) ручейники сходны с большинством чешуекрылых. Однако у примитивных представителей чешуекрылых имагинальный ротовой аппарат сохраняет грызущую специализацию с действующими мандибулами, что свидетельствует о независимой редукции имагинальных мандибул у ручейников и у хоботных бабочек [см. Amphiesmenoptera (12)]. Сосущая специализация ротового аппарата у них также возникла независимо: исходный хоботок бабочек образован удлиненными максиллярными галеами [ср.: Glossolepidoptera (1)], тогда как исходный хоботок ручейников образован увеличенным гипофаринксом.
В отличие от имаго, у личинок ручейников ротовой аппарат сохраняет исходную грызущую функцию и развитые мандибулы [см. Amphiesmenoptera (12)].
Плезиоморфии и разнообразие Trichoptera
(5) Крылья. Крыловой аппарат бывает бимоторным или переднемоторным. Примитивные ручейники бимоторные: их передние и задние крылья почти одинаково хорошо развиты, сходны по форме и жилкованию, и в полете не сцеплены. В некоторых таксонах ручейников независимо друг от друга произошел переход к переднемоторности, при которой в полете заднее крыло сцеплено с передним, а переднее крыло имеет выраженный торнус [см. Pterygota (4F)]. В этих случаях заднее крыло лишь немного короче переднего, так что торнус переднего крыла находится в дистальной части крыла. У некоторых переднемоторных ручейников переднее крыло узкое, с прямым задним краем, а заднее крыло сильно расширено за счет увеличенной инверталярной области, которая в этом случае приобретает способность складываться веерообразно [см. Neoptera (1a)]. Сцепка крыльев бывает амплексиформной (благодаря простому налеганию заднего края переднего крыла на передний край заднего) или обеспечивается специализированными продольными рядами щетинок на вентральной стороне переднего крыла около его заднего края и/или на дорсальной стороне заднего крыла около его переднего края. В разных таксонах ручейников сцепочные щетинки имеют разную форму и расположение – от прямых и направленных дистально, до сильно изогнутых и направленных поперечно (Stocks 2010a, 2010b).
У некоторых ручейников крылья вестигиальные или утрачены; у некоторых видов самка короткокрылая или бескрылая, тогда как самец крылатый (то же у многих других насекомых – см. Указатель признаков [2.99]).
Размеры. Длина тела 1,5–45 мм.
Геологический возраст. Юра – ныне (Rasnitsyn & Quicke 2002). По жилкованию крыльев Trichoptera не отличаются от других Amphiesmenoptera, поэтому некоторые авторы указывают более древней возраст Trichoptera, вплоть до пермского (см. Amphiesmenoptera).
Состав и распространение. Около 15 000 видов; всесветно (Zhang 2011).
Статус Trichoptera. Если рассматривать только современных насекомых, то таксон Trichoptera представляется бесспорно голофилетическим. Однако некоторые авторы относят к Trichoptera таких вымерших представителей Amphiesmenoptera, которые не имеют ни аутапоморфий Lepidoptera, ни аутапоморфий Trichoptera (см. Amphiesmenoptera: «Классификации»); включение таких насекомых в состав Trichoptera делает таксон Trichoptera плезиоморфоном. Употребление названия «Trichoptera» для этого плезиоморфона не противоречит правилам циркумскриптной номенклатуры, поскольку исходно это название выло опубликовано без учета вымерших таксонов (Kirby 1813). Однако целесообразно ограничить таксон Trichoptera таким образом, чтобы он оставался голофилетическим (см. раздел I.5.2.1 и рис. 1.3).
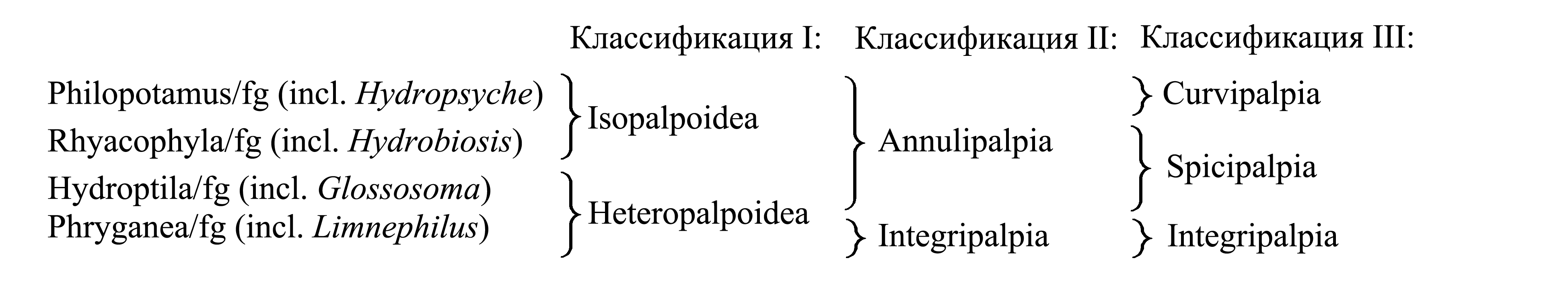
Классификации Trichoptera
I. Деление Trichoptera на Isopalpoidea и Heteropalpoidea.
1. Isopalpoidea Kolenati 1848 {syn.circ.: Aequipalpidae Kolenati 1859; nom.typ: Philopotamus/fg [fg:1829] (incl. Rhyacophila)}. Личинки с прогнатной головой и длинными анальными ложноножками [см. Trichoptera (2)], не строят переносных домиков. Личинки либо обитают открыто, либо используют естественные скважины, либо строят стационарные убежища – галереи или ловчие сети.
2. Heteropalpoidea Kolenati 1848 {syn.circ.: Inaequipalpidae Kolenati 1859; nom.typ.: Phryganea/fg [f:1804; g:1758] (incl. Hydroptila). Оба названия связаны с тем, что у самцов число члеников максиллярных щупиков сокращено до 4 или 3, тогда как у самки сохраняются все 5 члеников (Kolenati 1848). Личинки с гипогнатной головой и без анальных ложноножек, строят переносные домики.
II. Деление Trichoptera на Annulipalpia и Integripalpia.
1. Annulipalpia Martynov 1924d {syn.circ.: Vericloacia Weaver 1984; nom.typ: Philopotamus/fg [fg:1829] (incl. Rhyacophila, Hydroptila)}. Название «Annulipalpia» связано с тем, что у части представителей щупики имаго имеют кольчатый последний членик (см. III: Curvipalpia).
2. Integripalpia Martynov 1924d {syn.circ.: Dicloacia Weaver 1984; nom.typ.: Phryganea/fg [f:1804; g:1758]}.
III. Деление Trichoptera на Curvipalpia, Spicipalpia и Integripalpia.
1. Curvipalpia Weaver 1984 {nom.typ.: Philopotamus/fg [fg:1829]}. Название связано с тем, что у имаго последний членик лабиального и максиллярного щупика [т.е. 3-й членик лабиального щупика и 5-й членик максиллярного щупика – см. Trichoptera (4)] удлинен, гибкий, подразделен на вторичные кольца (у некоторых видов щупики редуцированы, и этот признак не выражен).
2. Spicipalpia Weaver 1984 {nom.typ.: Hydroptila/fg [f:1836; g:1819] (incl. Rhyacophila)}.
3. Integripalpia (см. II).
* * *