КЛАДОЭНДЕЗИС НАСЕКОМЫХ

![]()
|
КЛАДОЭНДЕЗИС НАСЕКОМЫХ |
|||||||
|
|
|
fg | |||||
VI-1.1.1. Diplura Börner 1904 – двухвостки
|
Названия, однозначно соответствующие по циркумскрипции: — Endotropha = Entotrophi Grassi 1888; — Archinsecta Haeckel 1896; — Diplura Börner 1904a; — Rhabduradelphia Crampton 1916a, 1916b; — Cryptognatha Krausse & Wolff 1919; — Dicelluradelphia = Pandicellura Crampton 1924; — Entotrophica Tillyard 1926; — Entotrophida Pearce 1936; — Diplurata Bey-Bienko 1962; — Eometabola Chen 1962; — Polyentomata Grandi 1966. По циркумскрипции соответствует также таксонам под типифицированными названиями, образованными от родовых названий Iapyx и Campodea: Iapygidae, Campodeidae, Campodina, Campodeoidea и др. Иерархическое название: Iapyx/f1=Campodea/g1 [f:1864; g:1842]. |
|
Ранг. Со второй половины XX века – обычно от отряда до класса; в конце XIX и начале XX века – часто подотряд; ранее – семейство.
Название. Название Diplura является общепринятым названием для этого таксона, хотя имеются более старые названия – Endotropha и Archinsecta.
Аутапоморфии Diplura. Все двухвостки имеют однообразную форму тела с прогнатной головой и удлиненным гибким туловищем. О голофилии этого таксона свидетельствуют уникальные апоморфии (1) и (2).
(1) Строение мандибул. Мандибулы [находящиеся в челюстных карманах – см. Entognatha (1)] однообразного строения: вытянуты вдоль оси причленения, без молы (в отличие от некоторых Collembola), инцизор всегда четырехзубчатый (рис. 6.3.1F)
Помимо инцизора на мандибуле Diplura бывает развита лишь простека; она имеется у Rhabdura и Prodicellura (рис. 6.3.3D), но отсутствует у Dicellurata. Некоторые авторы гомологизируют эту простеку с кинетодонтом [см. Mandibulata (1a)] и на основании этого считают наличие простеки плезиоморфией. Было ошибочно описано, что простека приводится в движение особой мышцей (Tuxen 1959, Manton 1964); наличие этой мышцы расценивалось как свидетельство того, что простека является члеником конечности и имеет древнее происхождение [об ошибочной гипотезе о первично двучлениковом строении мандибулы – см. Mandibulata (1)]. В действительности у Mandibulata ни кинетодонт (имеющий древнее происхождение), ни простека (которая может быть новообразованием) не имеют собственных мышц. За мышцу простеки у Diplura принимали задний дорсальный аддуктор мандибулы (рис. 6.3.1F: PDm); у других насекомых эта мышца так же всегда имеется и бывает самой мощной среди мандибулярных мышц [см. Mandibulata (1)], но у Diplura, в связи с тем, что эта мышца направлена почти параллельно оси вращения мандибулы, она ослаблена и бывает очень тонкой.
(2) Сцепка максилл и суперлингв. Максиллы сцеплены с суперлингвами благодаря тому, что каждая суперлингва имеет на латеральной стороне склеротизованный выступ, который входит в выемку на вентральной стороне максиллярной галеи (рис. 6.3.1E). У других насекомых, многоножек и ракообразных, имеющих суперлингвы [см. Mandibulata (3)], подобная сцепка отсутствует.
(3) Редукция глаз. Глаза [исходно вестигиальные – см. Entognatha (2)] полностью утрачены. То же у многих других насекомых (см. Указатель признаков [1.20] и табл. A).
Признаки Diplura неясного филогенетического значения
(4) Стернальные аподемы. Стерниты груди могут иметь непарные аподемы – спины. Спина (spina) представляет собой эктодермальную аподему, выстланную склеротизованной кутикулой; она отходит от вентральной стенки тела дорсально и/или назад, и проходит между коннективами нервной цепочки [см. Metameria (2)]; ее вершина, выступающая дорсальнее нервной цепочки, служит местом прикрепления мышц, проходящих дорсальнее нервной цепочки; эти мышцы идут к основаниям ног соответствующего сегмента и к определенным стернитам туловища. У Dicellurata каждый из трех сегментов груди несет спину, являющуюся продолжением Y-образного склерита: передние парные ветви этого склерита служат сочленовными мыщелками тазиков ног, а задняя непарная ветвь продолжается внутрь тела в виде спины (рис. 6.3.1C, 6.3.2D). У других двухвосток спины менее развиты; у изученных мной представителей Rhabdura спина имеется только на переднегруди, а Y-образные склериты отсутствуют на всех сегментах. Возможно, спины двухвосток гомологичны спинам крылатых насекомых [ср.: Pterygota (3)].
(5) Стерниты брюшка цельные. Стилигеральные швы [см. Hexapoda (7)] отсутствуют на всех сегментах брюшка (уромерах) (в отличие от многих Triplura), так что стерностили и кинетапофизы [отсутствующие на генитальных уромерах – см. (7)] если имеются, то сидят на заднем крае цельного стернита. Неясно, является ли это плезиоморфией, или аутапоморфией Diplura, или аутапоморфией Entognatha (поскольку у Entognatha-Ellipura нет ни швов, ни стерностилей, ни кинетапофизов исходного для Hexapoda строения).
В отличие от стилигальных швов, стерностили и кинетапофизы могут присутствовать на прегенитальных (I–VII) уромерах. Стерностили и подходящие к ним мышцы имеются либо на всех I–VII уромерах (у Prodicellura и Dicellurata), либо на II–VII уромерах (у Rhabdura). У Rhabdura и Prodicellura каждый стерностиль несет апикальный шип [см. Hexapoda (7)]; у Dicellurata апикальные шипы отсутствуют. Кинетапофизы первой пары у Rhabdura и Prodicellura невтягивающиеся [см. ниже, Rhabdura (1) и Prodicellura (2)] (рис. 6.3.3С); у Dicellurata они либо являются втяжными пузырьками, либо утрачены. Кинетапофизы II–VII пар либо являются втяжными пузырьками, либо утрачены (табл. VI.2).
(6) Положение полового отверстия. Половое отверстие самца и самки непарное (в отличие от самцов Protura), расположено между VIII и IX стернитами брюшка (рис. 6.3.2A, В; 6.3.3A) (то же у Protura); возможно, это исходное состояние для скрыточелюстных [см. Entognatha (4)].
(7) Отсутствие придатков VIII и IX уромеров. На генитальных сегментах брюшка [т.е. на VIII и IX уромерах – см. (6)] стерностили и кинетапофизы всегда отсутствуют и у самца, и самки [при этом они имеются на прегенитальных уромерах – см. (5)] (рис. 6.3.2A, В; 6.3.3A). Поскольку именно кинетапофизы VIII и IX уромеров образуют яйцеклад самки у Amyocerata (рис. 6.6.3), можно предположить, что отсутствие такого яйцеклада у самок Diplura и других Entognatha является результатом редукции [см. Hexapoda (13)]. Копулятивный аппарат, свойственный Protura, также отсутствует. Обычно около полового отверстия нет никаких образований, лишь у самцов некоторых Dicellurata имеется пара небольших придатков по бокам от полового отверстия. Оплодотворение сперматофорное [подробно описано для представителей Rhabdura (Bareth 1961)].
(8) Отсутствие трахеостий VIII уромера. На брюшке имеется не более 7 пар трахеостий – по паре на I–VII уромерах; в отличие от многих Amyocerata, трахеостии VIII уромера отсутствуют. У Rhabdura трахеостии брюшка совсем утрачены. У других Entognatha абдоминальные трахеостии также утрачены, поэтому неясно, является ли редукция трахеостий VIII уромера аутапоморфией Diplura.
В отличие от абдоминальных, грудные трахеостии всегда имеются и служат дыхальцами; среди всех насекомых только у Diplura встречается полный набор грудных дыхалец [две пары сегментарных и две пары межсегментарных – см. Hexapoda (4b) и (9)]. Все 4 пары грудных дыхалец имеются у Dicellurata (рис. 6.3.1A) и Prodicellura-Octostigma/fg; у других Prodicellura и у Rhabdura число пар грудных дыхалец сокращено до 3 или 2 (табл. VI.2).
(9) Строение X уромера. Склеротизованная область десятого уромера [последнего – см. Hexapoda (8)] представляет собой цельное кольцо, не разделенное на тергит и стернит (в отличие от предыдущих I–IX уромеров, у которых тергит и стернит ясно разделены парой продольных латеральных швов); десятый уромер всегда несет пару подвижных придатков – псевдокаудалий (pseudocaudalii), причлененных к нему сзади, по бокам от анального отверстия; между основаниями псевдокаудалий бывает развит эпипрокт (epiproctus) – небольшой отчлененный выступ, расположенный дорсальнее анального отверстия. Эпипрокт развит у Rhabdura и Prodicellura, но не выражен у Dicellurata. К основаниям псевдокаудалий подходят мышцы, неподвижные основания которых крепятся к стенкам десятого уромера: мышцы, раздвигающие псевдокаудалии, крепятся к латеральным стенкам, а мышцы, сдвигающие псевдокаудалии, крепятся к вентральной стенке, медиальнее пары продольных стернальных мышц (рис. 6.3.2A–B). Вероятно, исходно с парой псевдокаудалий связана пара отверстий протоков желез, расположенных в задней части брюшка: у всех Prodicellura отверстия желез открываются на вершинах псевдокаудалий; у Procampodea/fg (в составе Rhabdura) и у Parajapyx/fg (в составе Dicellurata) отверстия желез открываются у оснований псевдокаудалий; у прочих Rhabdura и Dicellurata железы отсутствуют (табл. VI.2).
Псевдокаудалии двухвосток обычно называют «церками» по сходству с церками амиоцерат; однако они отличаются от церков тем, что их мышцы-аддукторы крепятся к участку стенки тела, соответствующему не тергиту, а стерниту [см. Hexapoda (8a)]. У Dicellurata псевдокаудалии превращены в пару мощных хватательных клещей, в связи с чем десятый уромер увеличен и склеротизован, а пара мышц, сдвигающих псевдокаудалии, крепятся не только к вентральной стенке десятого уромера, а также к его латеральной и дорсальной стенке, заполняя почти десятый уромер; при этом мышцы, раздвигающие псевдокаудалии, могут быть утрачены (рис. 6.3.2C).
Некоторые авторы необоснованно интерпретировали эпипрокт как вестигий XI сегмента брюшка, хотя в действительности он не является частью туловища, и в нем не проходит кишечный канал [см. Hexapoda (8b)]. Форма и функция псевдокаудалий различается у разных двухвосток (рис. 6.3.3B, F, G) [см. ниже, Rhabdura (4), Prodicellura (3) и Dicellurata (1)].
Неясно, является ли такое строение десятого уромера и наличие псевдокаудалий аутапоморфией Diplura или Entognatha в целом, поскольку у других Entognatha десятый уромер утрачен [см. ниже, Ellipura (3)].
(10) Предличинка. Из яйца выходит предличинка [см. Gnathopoda (3b)] – неподвижная стадия с неясно расчлененными придатками; после линьки она превращается в личинку I возраста, похожую на взрослое насекомое, с таким же как у взрослого насекомого числом члеников антенн [см. (11)] и с развитыми брюшными придатками [см. (5)]. Эта особенность известна лишь для некоторых видов Rhabdura и Dicellurata, для которых изучен онтогенез (Orelli 1956, Smith 1961). Стадия предличинки имеется и у некоторых других насекомых (см. Указатель признаков [5.4]).
Плезиоморфии и изменчивые признаки Diplura
(11) Антенны [т.е. антенны I – см. Atelocerata (1)] с определенным или немного варьирующим числом члеников, у разных видов имеют от 13 до 70 члеников (в отличие от 4 члеников у Collembola и отсутствия антенн у Protura); все членики, за исключением самых дистальных, имеют собственную мускулатуру (рис. 6.3.1B) (в отличие от Amyocerata); число члеников не меняется в постэмбриогенезе, лишь в старших возрастах число члеников может немного увеличиваться путем деления самого дистального членика (Imms 1940, Orelli 1956, Smith 1961) (в отличие от Symphyla и др., у которых число члеников увеличивается начиная с ранних возрастов путем деления проксимальных члеников).
(12) Оральные складки [см. Entognatha (1)] не сходятся на вентральной стороне головы, так что нижняя губа в проксимальной части имеет вид широкой полосы, а не вентрального желоба (в отличие от Ellipura); фулькрум сохраняет U-образную форму (в отличие от Y-образной у Protura) (рис. 6.2D–E). Лабиальные щупики сохраняются у Dicellurata и некоторых Prodicellura; у Rhabdura и некоторых Prodicellura утрачены (табл. VI.2).
(13) Претарсус с парой коготков (в отличие от Ellipura), часто между ними имеется первичный непарный коготок – дактилоподит (рис. 6.3.1D) [см. Hexapoda (6)].
(14) Сохраняются линьки в половозрелой стадии (среди Hexapoda то же у Collembola и Triplura, в отличие от Protura и Pterygota). Общее число возрастов бывает большим, порядка 30.
Геологический возраст. Мел – ныне. Из нижнего мела Бразилии описан представитель Dicellurata – Ferrojapyx vivax Wilson & Martill 2001. Из палеогенового балтийского янтаря и более поздних отложений описаны представители Rhabdura и Dicellurata. На основании архаичного строения Diplura и того, что Entognatha известны начиная с девона, можно предполагать, что Diplura существовали еще в девоне. Однако из-за мелких размеров и нежных покровов большинства представителей они имеют мало шансов сохраниться в ископаемом состоянии.
Помимо насекомых, бесспорно относящихся к Diplura, к этому таксону были отнесены Osellia articulicornis Olfers 1907 из балтийского янтаря и Testjapyx thomasi Kukalova-Peck 1987 из верхнего карбона США. Эти насекомые, по мнению описавших их авторов, совмещают в себе признаки Diplura и Amyocerata, на основании чего делаются далеко идущие выводы о филогении насекомых. Однако на самом деле Osellia articulicornis является молодой личинкой уховертки (Dermatoptera) (Silvestri 1912). Testjapyx thomasi имеет 8 сегментов брюшка с парой клещевидных придатков на последнем из них (Kukalova-Peck 1987); это свойственно Dermatoptera, тогда как у всех Diplura (в том числе у Dicellurata, имеющих клещевидные придатки), брюшко состоит из 10 хорошо видимых сегментов.
Систематическое положение Diplura. Помимо принятого здесь отнесения Diplura к Entognatha предлагались следующие классификации.
I. Объединение Diplura и Triplura в таксон Cinura Packard 1883 {synn.circ.: Thysanuroidea Escherich 1914 = Eustyligera Crampton 1916b = Propleomerentoma Krausse & Wolff 1919 = Panthysanura Crampton 1920 = Mesapterygota Crampton 1924 = Styligera Crampton 1938; по циркумскрипции также соответствует: «Thysanura» sensu Lubbock 1873 (non Leach 1815a)}. Это объединение основано на общем признаке – наличие стерностилей и кинетапофизов на прегенитальных (I–VII) уромерах [см. Hexapoda (7)]. Судя по тому, что у Triplura и Pterygota имеются сериально гомологичные им придатки на генитальных (VIII–IX) уромерах, этот признак исходно присущ Amyocerata (см. II). и является исходным для насекомых [см. Hexapoda (7)], так что таксон Cinura является парафилетическим – предковым и для Ellipura, и для Pterygota.
II. Объединение Diplura и Amyocerata в таксон Pleomerentoma (см. Hexapoda: «Классификации» III).
III. Объединение Diplura, Collembola и Amyocerata в таксон Holomerentoma, противопоставляемый Protura (см. Protura: «Систематическое положение» I).
IV. Объединение Diplura и Protura в таксон Nonoculata (см. Entognatha: «Классификации» II).
Статус Diplura. В прошлом существовало мнение, что Diplura – наиболее примитивная группа среди насекомых (Lubbock 1874, Haeckel 1896). При этом усматривали сходство между Diplura (относимыми к роду Campodea) и личинками некоторых Pterygota, в связи с чем эти личинки получили название «камподиевидные» («Campodea-form» по Lubbock 1874); это сходство расценивали как результат действия «биогенетического закона», который выражается в проявлении предковых признаков на ранних стадиях онтогенеза (об ошибочности этого закона см. раздел I.4.2.1). В качестве общих примитивных признаков двухвосток и камподиевидными личинок разных крылатых насекомых – поденок, стрекоз и некоторых жуков – указывали наличие развитых антенн, ног и хвостовых придатков (Lubbock 1874: Pl. 3, 4); однако в действительности хвостовые придатки этих насекомых не гомологичны: у двухвосток это псевдокаудалии [см. Diplura (9)], у поденок это каудалии [ср.: Amyocerata (8)], у личинок стрекоз это придатки парапроктов и эпипрокта [ср.: Odonata (10)], а у личинок жуков это урогомфы, расположенные не на последнем, а на предпоследнем сегменте брюшка [ср.: Elytrophora (12)]. В качестве довода в пользу происхождения всех насекомых от двухвосток указывали также скрыточелюстной ротовой аппарат двухвосток, который считали промежуточным между грызущим и сосущим, исходным как для грызущего ротового аппарата большинства насекомых, так и для сосущего ротового аппарата хоботных. Однако из сравнения с другими членистоногими следует, что ротовые придатки насекомых происходят от конечностей, расположенных снаружи головы, а погружение мандибул и максилл в челюстные карманы является вторичным; при этом способ погружения мандибул и максилл в голову принципиально различен у двухвосток [см. Entognatha (1)] и у хоботных [ср.: Acercaria (1), Condylognatha (1b) и Arthroidignatha (1a)].
Diplura отличаются от общего предка Hexapoda, по крайней мере, наличием челюстных карманов [см. Entognatha (1)].
Некоторые авторы считают Diplura парафилетической группой, предковой для Ellipura; на основании этого высказывают сомнения в целесообразности существования таксона Diplura (Hennig 1981, Stys & Bilinski 1990). У разных представителей Diplura выявлено разное строение овариев [см. Hexapoda (12)]: у Rhabdura оварии не разделены на овариолы, у Anajapyx/fg каждый оварий разделен на 2 овариолы, а у Dicellurata каждый оварий разделен на 7 овариол. Высказываются разные точки зрения о филогенетическом значении этих признаков: согласно одной из них, наличие семи овариол считается плезиоморфией, а упрощенное строение овариев – синапоморфией Rhabdura и Ellipura (Stys & Bilinski 1990) [ср. Ellipura (4)]; согласно другой версии, наоборот, простое строение овариев считается плезиоморфией, а наличие семи овариол – синапоморфией Dicellurata и Amyocerata, объединяемых в таксон Ovariolata [см. Hexapoda (12)]. Обе эти версии основаны только на данных о строении овариев у единичных видов и не подтверждаются другими признаками. Следует упомянуть, что в некоторых явно голофилетических таксонах насекомых число овариол сильно варьирует: например у различных представителей Diptera бывает одна овариола (как у Rhabdura), две овариолы (как у Anajapyx/fg), 5–7 овариол (как у Dicellurata) и большее число овариол, достигающее сотню.
Единообразное строение мандибул [см. Diplura (1)] и наличие сцепки суперлингв с максиллами [см. Diplura (2)] свидетельствуют о голофилии Diplura.
Классификация Diplura. Таксон Diplura делят либо на Rhabdura s.l. (включая Prodicellura) и Dicellurata, либо на Rhabdura s.str. и Dicellura s.l. (включая Prodicellura). Некоторые признаки таксонов в составе Diplura показаны на табл. VI.2.
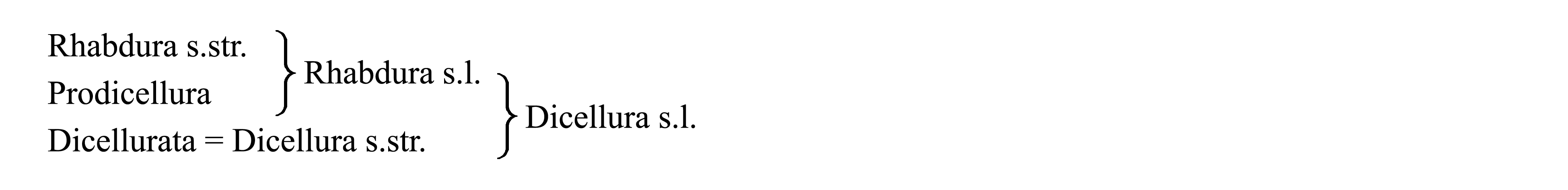
Здесь мы принимаем в составе Diplura три подчиненных таксона – Rhabdura s.str., Prodicellura (стр. 0000) и Dicellurata (стр. 0000).
Diplura, или Iapyx/f1=Campodea/g1
![]() 1. Rhabdura, или Campodea/f1=g2
1. Rhabdura, или Campodea/f1=g2
![]() 2. Prodicellura, или Projapyx/fg1
2. Prodicellura, или Projapyx/fg1
![]() 3. Dicellurata, или Iapyx/f2=g1
3. Dicellurata, или Iapyx/f2=g1