КЛАДОЭНДЕЗИС НАСЕКОМЫХ

![]()
|
КЛАДОЭНДЕЗИС НАСЕКОМЫХ |
|||||||
|
|
|
fg | |||||
IX-1.2.2.2. Arthroidignatha Spinola 1850 – членистохоботные
(Somobialia![]() Eucaryota
Eucaryota![]() Metazoa
Metazoa![]() Monostomata
Monostomata![]() Haemataria
Haemataria![]() Peritoneata
Peritoneata![]() Metameria
Metameria![]()
Gnathopoda![]() Euarthropoda
Euarthropoda![]() Mandibulata
Mandibulata![]() Atelocerata
Atelocerata![]() Hexapoda
Hexapoda![]() Amyocerata
Amyocerata![]()
Pterygota![]() Metapterygota
Metapterygota![]() Neoptera
Neoptera![]() Eumetabola
Eumetabola![]() Parametabola
Parametabola![]() Acercaria
Acercaria![]() Condylognatha
Condylognatha![]() Arthroidignatha)
Arthroidignatha)
(рис. 9.6.1–9.12.4)
9.6.1, 9.7.1,
9.7.2
|
Названия, однозначно соответствующие по циркумскрипции: — Semivaginata Schluga 1767; — Arthroidignatha Spinola 1850; — Hemipteroidea Handlirsch 1903; — Hemipteradelphia = Panhemiptera Crampton 1924; — Hemipteriforma Metcalf 1951; — Hemipteroida Müller 1963; — Hemipterida Boudreaux 1979b; — Homopteroidea Brodsky 1994. По циркумскрипции также соответствует: — «Hemiptera» sensu Geoffroy 1762 (non Linnaeus 1758); — «Ryngota» auct. (non Fabricius 1775); — «Rhyngota» auct. (non Fabricius 1803); — «Rhynchota» auct. (non Burmeister 1835; nec Rhynchota Billberg 1820); — таксонам под типифицированными названиями, образованными от родового названия Cimex: Cimicida, Cimicidea и др. Иерархическое название: Cimex/f2=Cicada/g4 [f:1781(ord.3); g:L.1758(gen.195)] (sine Thrips; incl. Aphis). |
|
Ранг. Обычно отряд или надотряд.
Названия. Иногда этому таксону приписывают название Ryngota Fabricius 1775 или его измененное написание Rhyngota Fabricius 1799. В действительности оба эти названия не соответствуют по своей исходной циркумскрипции рассматриваемому таксону, так как Фабрициус относил к Ryngota, помимо прочего, роды Pulex и Thrips (т.е. таксоны, называемые ныне Aphaniptera и Thysanoptera).
Широко признанным названием этого таксона является Rhynchota (измененное Rhyngota), однако и оно не соответствует ему по исходной циркумскрипции. Исходно название Rhynchota Burmeister 1835 относилось к таксону, включающему, помимо прочего, вшей (см. Siphunculata: «Систематическое положение» I). Кроме того, это название является преоккупированным: ранее такое же название Rhynchota Billberg 1820 давалось для блох (Aphaniptera).
Часто этот таксон называют также Hemiptera. Старинное название Hemiptera в XVIII веке давалось самым различным таксонам: сборной группе, включавшей скорпионов, клопов, светляков, прямокрылых и муравьев (Linnaeus 1735); группе, объединяющей насекомых с умеренно жесткими передними крыльями – хоботных и прямокрылых в широком смысле (Linnaeus 1789), только прямокрылых в широком смысле (Retzius 1783) или только хоботных в широком смысле. В 10-м издании «Systema Naturae» (Linnaeus 1758) (являющемся исходным для приоритета зоологических названий – см. I.6.3.1.2) название Hemiptera отнесено к группе, включавшей не только членистохоботных, но и трипсов, т.е. по циркумскрипции соответствующей Condylognatha. В таком значении это название широко употреблялось в течение всего XIX века; некоторые авторы середины и конца XIX века относили к Hemiptera, помимо членистохоботных и трипсов, также вшей. В XX веке название Hemiptera в этих значениях почти не используется, и чаще всего употребляется в одном из двух сильно различающихся значений: либо для Arthroidignatha, либо для Heteroptera. Таким образом, во избежание путаницы желательно вообще не использовать название «Hemiptera».
Название Arthroidignatha Spinola 1850 однозначно соответствует по циркумскрипции данному таксону; оно было предложено автором, установившем этот таксон и давшем ему правильную характеристику.
Более раннее забытое название Semivaginata Schluga 1767 является латинским эквивалентом греческого названия «Hemelytrata», т.е. отражает признак, свойственный лишь подчиненному таксону (см. ниже, Hemelytrata); употребление этого названия может вызвать такие же недоразумения, как употребление сходного по значению названия «Hemiptera».
Аутапоморфии Arthroidignatha
(1) Хоботок. Колюще-сосущий ротовой аппарат [см. Condylognatha (1)] усложнен (рис. 9.6.1, 9.7.1A, 9.7.2): в состав внутреннего хоботка (proboscis interior), помимо стилетов максиллярных лациний, видоизмененных особым образом [см. (1b)], вошли видоизмененные мандибулярные стилеты [см. (1a)]; внутренний хоботок заключен во внешний хоботок (proboscis exterior), образованный сильно видоизмененной нижней губой [см. (1d)]; слюнной шприц имеет особое строение [см. (1c)]. В таком аппарате весь внутренний хоботок работает как единая конструкция; он в совершенстве приспособлен для прокалывания покровов и высасывания жидкой пищи, которой исходно является сок растений [см. (1e)]. Со строением ротового аппарата связаны общие особенности эволюции, проявляющиеся в разных таксонах членистохоботных [см. (1f)].
Поскольку стилеты внутреннего хоботка скрыты внутри внешнего хоботка, а щупики утрачены [см. (1b) и (1d)], снаружи весь ротовой аппарат выглядит как цельный непарный членистый внешний хоботок (с чем связано название Arthroidignatha); лишь спереди к основанию хоботка прижата более короткая верхняя губа.
Ротовой аппарат принципиально одинаково устроен у имаго и у личинок всех возрастов [которых бывает от 2 до 5 – см. Condylognatha (3)]; лишь у непитающихся стадий ротовой аппарат может быть вестигиальным или утрачен [см. ниже, Gynaptera (1), Gallinsecta (1)].
Как и в характеристике Condylognatha, здесь в разделах (1a)–(1d) ротовые части условно описаны так, как они расположены при гипогнатном положении головы (когда ротовые придатки направлены вершинами вентрально, клипеус и верхняя губа находятся спереди, а нижняя губа сзади), хотя среди Arthroidignatha имеются также представители с опистогнатной и прогнатной головой.
(1 a) Модификация мандибул (рис. 9.6.1). Мандибула [скрытая в кармане и дистально вытянутая в стилет – см. Condylognatha (1a)] приобрела подвижный мандибулярный рычаг и мышцы-протракторы, идущие к особой мандибулярной пластинке головы, а мандибулярные стилеты сцеплены с максиллярными, образуя вместе с ними единый внутренний хоботок.
Из двух мыщелков мандибулы [см. Metapterygota (2)] сохраняется только один – исходно задне-латеральный. Имеется мандибулярный рычаг, который представляет собой обособленный палочковидный склеротизованный участок мандибулы, исходно являющийся проксимальным и несущий мыщелок. От мыщелка, соединяющего мандибулярный рычаг с краем головной капсулы, мандибулярный рычаг тянется перпендикулярно поверхности головной капсулы вглубь мандибулярного футляра и там сочленен с остальной частью мандибулы, которая представляет собой мандибулярный стилет. Рычаг и стилет мандибулы, подвижно соединенные под углом друг к другу, по строению и принципу действия напоминают рычаг и стилет максиллярной лацинии, хотя их происхождение совершенно различно [максиллярный рычаг, в отличие от мандибулярного, является внутренней аподемой – см. Condylognatha (1b)]. Подвижно сочлененный рычаг позволяет мандибуле вдвигаться и выдвигаться, совершая прокалывающие движения.
Благодаря такому строению мандибулы ее задняя дорсальная мышца [исходно являющиеся аддуктором – см. Mandibulata (1)], служат ретрактором. Эта мышца-ретрактор от своего основания разделяется на кранио-мандибулярную мышцу, сохраняющую исходное прикрепление к головной капсуле, и тенторио-мандибулярную мышцу, крепящуюся к передней руке тенториума (рис. 9.6.1A). У некоторых Plantisuga исходные кранио-мандибулярные мышцы утрачены, и ретракторами служат только тенторио-мандибулярные мышцы [см. ниже, Plantisuga (1)].
Помимо ретрактора (или ретракторов), мандибула имеет мышцу-протрактор, идущую от нее к мандибулярной пластинке. Пара мандибулярных пластинок, называемых также параклипеальными долями, или уздечками, или лорумами (lora) образует часть наружной стенки головы по бокам от клипеуса [обычно крупного – см. Acercaria (4)]. Спереди (или медиально) мандибулярная пластинка непосредственно граничит с латеральным краем клипеуса; от клипеуса [служащего местом прикрепления цибариальных мышц – см. Amyocerata (3)] мандибулярная пластинка обычно отделена бороздой; иногда эта борозда не выражена, и мандибулярные пластинки выглядят как латеральные части клипеуса. Задний край мандибулярной пластинки плотно смыкается с передним краем максиллярной пластинки [см. (1b)]; между ними имеется щель, ведущая в мандибулярный футляр [исходно эта щель разделяет клипеус и максиллярную пластинку – см. Condylognatha (1a)].
Мандибулярный стилет имеет желоб на внутренней (медиальной) стороне, благодаря чему пара мандибулярных стилетов плотно охватывает с боков трубку, образованную парой максиллярных стилетов [см. (1b)]; в результате этого мандибулярные и максиллярные стилеты образуют единый внутренний хоботок. Двигаясь поочередно с максиллярными стилетами, мандибулярные стилеты участвуют в прокалывании субстрата [см. (1e)]. У многих Heteropteroidea мандибулярные стилеты не только охватывают максиллярные, но и прочно сцепляются с ними с помощью продольных гребней и пазов, образующих скользящие соединения.
Происхождение мандибулярных пластинок и крепящихся к ним мышц-протракторов мандибул вызвало дискуссию. Вероятно, мандибулярная пластинка является вентральным разрастанием щеки – т.е. латеральной области краниума, которая исходно вмещает неподвижное основание передней дорсальной мышцы мандибулы; соответственно, передняя дорсальная мышца [исходно служащая абдуктором мандибулы – см. Mandibulata (1)] стала протрактором (Weber 1928, Емельянов 2002). Согласно другой версии, мандибулярная пластинка представляет собой латеральную часть гипофаринкса, которая необычным образом увеличилась и выступает наружу; при этом предполагается, что протрактор является вентральной мандибулярной мышцей [слабо развитой у других насекомых – см. Metapterygota (2)] (Snodgrass 1938b). По третьей версии мандибулярная пластинка образовалась из латеральной части клипеуса (Kramer 1950); при этом происхождение протрактора оказывается неясным. Высказывалась также четвертая версия, по которой мандибулярная пластинка является частью мандибулы (подобно тому, как сходная с ней максиллярная пластинка – часть максиллы); однако это не согласуется с исходным строением мандибулы Hexapoda, не имеющей внутренних мышц.
(1 b) Модификация максилл (рис. 9.6.1, 9.7.2D). Наружные части максилл [не имеющих разделения на кардо и стипес – см. Acercaria (1)] в основании слиты с головной капсулой и представляют собой пару максиллярных пластинок – неподвижных продолжений латеральных стенок головной капсулы. Соответственно, максиллярная пластинка служит местом прикрепления мышц, выдвигающие максиллярный стилет; к максиллярной пластинке или к отходящей от нее аподеме крепится максиллярный рычаг [см. Condylognatha (1b)]. Максиллярная пластинка отделена от лежащей впереди нее мандибулярной пластинки [см. (1a)] щелью, ведущей в карман, в котором расположены максиллярный и мандибулярный стилеты. Максиллярный щупик полностью утрачен, так что снаружи максиллярные пластинки утратили сходство с максиллами и кажутся участками головной капсулы.
Максиллярные стилеты [исходно образующие канал для проведения пищи и слюны – см. Condylognatha (1b)] имеют такую форму поперечного сечения, что, складываясь вместе, они вместо одного канала образуют два изолированных канала, по которым жидкости движутся в противоположных направлениях: передний (дорсальный) канал служит для проведения жидкой пищи в глотку; задний (вентральный) канал служит для проведения слюны, которая подается в него из отростка гипофаринкса [см. (1c)]. У Plantisuga слюнной канал образован желобом, имеющегося только на левом максиллярном стилете; у Hemelytrata он образован либо желобом на правом максиллярном стилете, либо желобами и на правом, и на левом стилетах (рис. 9.6.1B, 9.7.2C) (Brozek 2006). У различных членистохоботных слюнный канал бывает такой же ширины как пищевой, либо шире его, либо ýже.
Среди членистохоботных такое строение является очень стабильным; лишь у некоторых водомерок (Heteroptera-Ploteres) максиллярные стилеты вторично упрощены таким образом, что имеется не два, а лишь один канал, поочередно функционирующий то как слюнный, то как пищевой.
Высказана следующая гипотеза о том, каким образом в ходе эволюции могло скачкообразно возникнуть такое соединение максиллярных стилетов, при котором получаются два полностью разделенных канала, проводящие жидкости в противоположных направлениях (Емельянов 2002). У предков Arthroidignatha максиллярные стилеты при складывании образовывали единственный канал, поочередно служащий для проведения пищи и слюны (как у современных Thysanoptera), но при этом стилеты были уплощены и расширены в медиальной плоскости (уплощенная форма максиллярных стилетов свойственна современным Saltipedes и другим Plantisuga). Верхняя губа была скручена в трубку так, что ее края налегали друг на дуга; максиллярные стилеты, проходя через такую асимметричную трубку, скручивались несколько винтообразно, а именно левый стилет смещался назад, а правый – вперед. В результате этого две половинки канала, образованного максиллярными стилетами, оказались разделенными, и таким образом из одного канала сразу, без переходных состояний, получилось два изолированных канала. Отросток гипофаринкса, подающий слюну, оказался в более заднем (вентральном) из этих каналов, и этот канал стал служить только для проведения слюны.
Максиллярные стилеты входят в состав внутреннего хоботка вместе с охватывающими их мандибулярными стилетами [см. (1a)].
(1 c) Модификация гипофаринкса (рис. 9.7.2C–D). Гипофаринкс образует слюнной шприц, усложненный по сравнению с подобным образованием у Thysanoptera [см. Condylognatha (1c)]; этот шприц накачивает слюну в задний из двух каналов, образованных максиллярными стилетами [см. (1b)]. Латеральные края гипофаринкса [завернутые назад и обрамляющие слюнной желоб – см. Condylognatha (1c)] смыкаются сзади (вентрально) по средней линии так, что образуют непарный канал шприца; в глубине этого канала имеется расширение – камера шприца, в которую сзади открывается непарный проток слюнных желез. Камера имеет склеротизованные стенки, а ее проксимальная стенка образует поршень – обособленный склеротизованный выступ, плотно входящий в камеру. Мышцы гипофаринкса сильно увеличены, вееровидные; их неподвижные основания широкие, прикреплены к латеральным стенкам гипофаринкса, которые увеличенным, склеротизованы и глубоко вдаются в голову, образуя пару крыловидных аподем; эти аподемы могут оканчиваться свободно или соединены с аподемами максиллярных пластинок и/или задними руками тенториума. Подвижные концы мышц крепятся к непарному сухожилию, отходящему проксимально от поршня шприца. При сокращении мышц поршень шприца отводится, камера расширяется и слюна накачивается из слюнного протока в камеру; при расслаблении мышц поршень под действием упругости стенок камеры вдвигается в камеру, и слюна под давлением накачивается в слюнной канал, по которому идет до вершины гипофаринкса. Вершина гипофаринкса вставлена между максиллярными стилетами так, что входит в задний канал, образованный ими (рис. 9.7.2C).
(1 d) Модификация нижней губы (рис. 9.6.1, 9.7.2B). Нижняя губа [исходно упрощенная – см. Acercaria (3)] крайне видоизменена и превращена в членистый внешний хоботок (proboscis exterior), называемый также рострум (rostrum): она сильно удлинена, ее латеральные края завернуты вперед и смыкаются (но не срастаются) спереди по прямой продольной линии так, что вся нижняя губа образует трубку, окружающую внутренний хоботок [см. (1e)]; эта трубка подразделена поперечными швами на несколько подвижных члеников.
У многих Arthroidignatha внешний хоботок имеет непарную базальную аподему; эта аподема представляет собой продолжение передней вогнутой стенки внешнего хоботка, вдающееся вглубь тела. Исходно базальная аподема склеротизована и служит местом прикрепления мышц, двигающих внешний хоботок (рис. 9.7.2B, 9.10.1A).
Судя по всему, исходным для Arthroidignatha является 3-члениковый внешний хоботок (Емельянов 1987). Внешний хоботок имеет 3 членика у Scytinelytra (рис. 9.10.1A), у части Gallinsecta, у всех Auchenorrhyncha-Euhomoptera (рис. 9.7.2A–B), у примитивных Auchenorrhyncha-Subtericornes и у Coleorrhyncha (рис. 9.8.1D). У этих насекомых 1-й и 2-й членики хоботка четко разделены только на задней стороне, а 2-й и 3-й членики четко разделены со всех сторон. У некоторых Auchenorrhyncha-Subtericornes внешний хоботок 4-члениковый или 5-члениковый благодаря разделению исходного второго членика. У большинства клопов хоботок 4-члениковый благодаря наличию дополнительного членика, находящегося проксимальнее входа в базальную аподему [см. ниже, Heteroptera (3) и рис. 9.8.2A]. У тлей хоботок 4-члениковый благодаря разделению на два исходного третьего членика [см. ниже, Gynaptera (4) и рис. 9.11.2]. В некоторых случаях число члеников хоботка вторично уменьшено из-за слияния тех или иных члеников, иногда расчленение хоботка совсем утрачено. У листоблошек хоботок 2-члениковый [см. ниже, Saltipedes (2) и рис. 9.9.1A, 9.9.2A]. У некоторых Gallinsecta число члеников хоботка сокращено от трех до 2 или 1. У некоторых клопов число члеников сокращено от исходных четырех до 3 или 1.
Во внешнем хоботке отсутствуют парные придатки, характерные для нижней губы других насекомых – глоссы, параглоссы и лабиальные щупики; так что неясно, каким частям исходной нижней губы гомологичны те или иные части внешнего хоботка. Предполагают, что первый членик образовался из постментума, второй – из прементума, а третий – из сросшихся глосс и параглосс (Емельянов, 2002). Согласно другой гипотезе, внешний хоботок образован не самой нижней губой, а сросшимися лабиальными щупиками [исходно 3-члениковыми – см. Hexapoda (3c)].
(1 e) Функционирование хоботка. Исходно хоботок приспособлен для питания соками и клеточным содержимым сосудистых растений: фитофагия свойственна всем Plantisuga, Auchenorrhyncha, Coleorrhyncha и части Heteroptera; лишь среди Heteroptera, помимо фитофагов, имеются факультативные и облигатные хищники, высасывающие гемолимфу из пойманных насекомых; несколько видов Heteroptera приспособились к сосанию кровью позвоночных животных. С питанием соками растений связаны модификации кишечника [см. (3)].
Все функции хоботка – и прокалывание покровов, и проведение жидкой пищи из субстрата в глотку, и проведение слюны из саливария в субстрат – осуществляются внутренним хоботком, который состоит из сложенных вместе или сцепленных парных мандибулярных и максиллярных стилетов [см. (1a) и (1b)].
Внутренний хоботок проходит внутри желоба внешнего хоботка [образованного нижней губой – см. (1d)]. Спереди к основанию внешнего хоботка прижата более короткая верхняя губа; иногда внутренний хоботок проходит в желобе на задней стороне верхней губы [см. Condylognatha (1d)], а дистальнее входит в желоб на передней стороне внешнего хоботка. В покое стилеты внутреннего хоботка не выступают за вершину внешнего хоботка и не видны снаружи; при питании насекомое приставляет вершину внешнего хоботка к субстрату, затем стилеты, скользя друг вдоль друга, по очереди выдвигается из вершины внешнего хоботка, и, благодаря этому, внутренний хоботок постепенно проникает в субстрат.
Внутренний хоботок может быть умеренной длины, так что во втянутом состоянии он остается прямым и помещается в голове и внешнем хоботке. Но у некоторых фитофагов, имеющих мелкие размеры и питающихся соками из глубоко лежащих тканей растения, внутренний хоботок удлинен настолько, что во втянутом состоянии образует петли той или иной формы, которые помещаются либо снаружи головы (у личинок Saltipedes), либо внутри кармана того или иного строения, называемого крумена (crumena) (у имаго Saltipedes, личинок и самок Gallinsecta, личинок Scytinelytra и у некоторых Heteroptera). Внутренний хоботок образует петли как единое целое, а составляющие его четыре стилета могут скользить друг вдоль друга, не раздвигаясь. Поэтому, когда внутренний хоботок сложен в виде петли, его выдвигание осуществляется следующим образом: сначала, благодаря поочередному сокращению протракторов и скольжению стилетов по петле, стилеты поочередно выдвигаются, но петля сохраняет свою форму и размер; затем весь внутренний хоботок зажимается в желобе внешнего хоботка дистальнее петли, а ретракторы стилетов, сокращаясь все одновременно, втягивают основание внутреннего хоботка и тем самым уменьшают изгиб его петли; затем цикл повторяется, пока петля не выпрямится полностью (Weber 1933).
(1 f) Особенности эволюции Arthroidignatha, связанные с наличием хоботка. В связи с приобретением колюще-сосущего хоботка, в совершенстве приспособленного для сосания сока из живых растений, у Arthroidignatha появились новые возможности в эволюции образа жизни. В то время как большинство наземных животных для добывания пищи вынуждены постоянно перемещаться, сосущие фитофаги могут добывать пищу, не перемещаясь. Членистохоботное небольших размеров может, внедрив свой внутренний хоботок в ткани растения, достичь им проводящего сосуда так, чтобы поступающего по сосуду сока было достаточно для полного обеспечения насекомого пищей. В этом случае насекомому выгоднее оставаться на одном месте и питаться из одного и того же сосуда, чем перемещаться и искать подходящий сосуд заново. В связи с этим следи членистохоботных, помимо обычных активно перемещающихся форм (Heteropteroidea, многие Auchenorrhyncha и др.) имеются малоподвижные и совсем неподвижные формы (личинки Scytinelytra, личинки и самки многих Gallinsecta и др.).
Другой особенностью эволюции членистохоботных является приобретение фильтрационной камеры [см. (3)]; благодаря ей насекомое, эффективно усваивая белки, содержащиеся в соке растения, выделяет из анального отверстия много воды с сахарами. Эти сахаристые экскременты могут пачкать растения, приводя к их угнетению и тем самым лишая насекомое возможности питаться, оставаясь на одном месте. Большую помощь малоподвижным и неподвижным членистохоботным в избавлении от жидких сахаристых экскрементов оказывают насекомые, поедающие эти экскременты. Это преимущественно имаго различных представителей Metabola (насекомых с полным превращением) – двукрылые [см. Diptera (1e)], ручейники [см. Trichoptera (4)], бабочки (Lepidoptera), перепончатокрылые (Hymenoptera) и др. В их питании белки не обязательны [поскольку имаго не растут – см. Pterygota (4C)], а углеводы важны для пополнения энергетических затрат. В некоторых случаях симбиоз Arthroidignatha с имаго Metabola привел к появлению особых коадаптаций; в частности, многие Plantisuga и некоторые личинки Auchenorrhyncha имеют поведенческие и морфологические приспособления к кормлению своими экскрементами рабочих муравьев.
(2) Слияние ганглиев. Абдоминальный нервный синганглий [см. Parametabola (6)] слит с заднегрудным ганглием. Как правило (за редким исключением) они слиты также со среднегрудным ганглием, в этом случае вентральная нервная цепочка состоит из 3 ганглиев – подглоточного синганглия, переднегрудного ганглия и заднего синганглия, объединяющего все прочие; в отдельных случаях переднегрудной ганглий слит с подглоточным и/или с задним синганглием (Pflugfelder 1937). Неуникальная апоморфия.
Признаки Arthroidignatha неясного филогенетического значения
(3) Фильтрационная камера. У некоторых членистохоботных, питающихся соками растений [см. (1)], кишечник имеет фильтрационную камеру. Поскольку в соках растений содержится большое количество воды и сахаров при относительно малом содержании белков (необходимых для развития насекомого, которое на всех стадиях своего жизненного цикла питается только соком одного вида растения), фильтрационная камера важна для удаления из кишечника избытка воды и сахаров. Фильтрационная камера образована тесным соединением двух удаленных участков кишечника, в результате чего образуется петля. Избыток воды и сахаров, имеющийся в пище, кратчайшим путем через соединение стенок кишечной петли выводится из организма, в то время как белковые и жировые компоненты усваиваются на более длинном пути в петле кишечника. В результате этого некоторые питающиеся соками членистохоботные выделяют из анального отверстия обильные жидкие сладкие выделения, известные как падь, или медвяная роса [о последствиях выделения пади см. (1f)].
Строение фильтрационной камеры различается в разных группах Arthroidignatha. Исходно фильтрационная камера образована соединением двух участков средней кишки [энтодермальной – см. Haemataria (1)] так, что пища в этих участках движется в противоположных направлениях; при этом передний участок кишки более широкий, а задний либо погружен в продольную складку переднего (у Scytinelytra и многих Auchenorrhyncha), либо петлями изгибается по его поверхности (у Saltipedes и некоторых Auchenorrhyncha). У Gallinsecta и Gynaptera в состав фильтрационной камеры включена также задняя, или прямая, кишка [эктодермальная – см. Haemataria (1)]: на задней кишке имеется вырост, охватывающий либо первичную фильтрационную камеру, либо только передний участок средней кишки (в этом случае первичное соединение двух участков средней кишки утрачено).
Фильтрационная камера имеется у многих Arthroidignatha, традиционно относимых к Homopterida (см. «Классификации» I) – у Saltipedes, Scytinelytra, у большинства Gallinsecta, у значительной части Gynaptera, у большинства Auchenorrhyncha. В то же время фильтрационная камера отсутствует у многих Gynaptera и др., перешедших к питанию клеточным соком (который, в отличие от сока из проводящей системы растения, содержит меньше воды и сахаров); фильтрационная камера отсутствует также у Auchenorrhyncha-Subtericornes и у всех Heteropteroidea.
Питание растительными соками является исходным типом питания Arthroidignatha [см. (1)], и лишь некоторые Heteroptera перешли к хищничеству и кровососанию. Поэтому возможно, что фильтрационная камера возникла у общего предка Arthroidignatha. Однако высказывается мнение о независимом возникновении фильтрационной камеры у разных Arthroidignatha.
В то время как в большинстве случаев сохраняется сквозной кишечник, у некоторых питающихся соками Gallinsecta и Heteroptera проход из энтодермальной средней кишки в эктодермальную заднюю кишку отсутствует, так что средняя и задняя кишка соединены лишь сплошным тяжем, не имеющим просвета (Goodchild 1963).
(4) Телотрофический яичник. Яичник [мероистический – см. Eumetabola (6)] телотрофического типа – т.е. трофоциты сконцентрированы в апикальной части овариолы (в трофарии), а от каждой яйцеклетки тянется питающий тяж к трофоцитам. В каждом кластере, получающемся при делении одного цистобласта, один или более цистоцитов развивается в яйцеклетки, а остальные цистоциты дают трофоциты (в отличие от политрофического яичника Panpsocoptera и многих Metabola, где в каждом кластере имеется лишь одна яйцеклетка и в отличие от паноистического яичника Thysanoptera, где все цистоциты образуют яйцеклетки) (Büning 1996). Неуникальная апоморфия, то же у Ephemeroptera и некоторых Metabola [см. Hexapoda (12)]. У некоторых членистохоботных (в частности, у некоторых Gynaptera) яичник не имеет трофоцитов, т.е. является паноистическим.
Геологический возраст. Пермь – ныне (Rasnitsyn & Quicke 2002).
Состав и распространение. Более 100 000 видов (Zhang 2011); всесветно.
Систематическое положение Arthroidignatha. Помимо принятого здесь объединения Arthroidignatha и Thysanoptera в таксон Condylognatha, некоторые авторы XVIII–XIX веков объединяли Arthroidignatha с теми или иными насекомыми, имеющими сосущий хоботок (вшами, бабочками, двукрылыми и/или блохами) в таксоны Ryngota Fabricius 1775, Haustellata Clairville 1798, Siphonostoma Latreille 1806, Rhynchota Burmeister 1835, Antliostomata Westwood 1839, Ectramoglossata Spinola 1850, Sugentia Haeckel 1866 и Pungentia Haeckel 1896 (см. «Название» и Pterygota: «Классификации» I). Эти таксоны были полифилетическими, поскольку сосущие хоботки независимо возникали в разных группах насекомых за счет специализации разных частей исходного грызущего ротового аппарата (см. Указатель признаков [1.36]).
Классификации Arthroidignatha. Некоторые авторы выделяют в составе Arthroidignatha непосредственно подчиненные таксоны Saltipedes, Scytinelytra, Gynaptera, Gallinsecta, Auchenorrhyncha, Coleorrhyncha и Heteroptera; иногда выделяют большее число непосредственно подчиненных таксонов, рассматривая вместо Auchenorrhyncha от 2 до 4 таксонов того же ранга. Предлагались следующие варианты деления Arthroidignatha на 2 подчиненных таксона.
I. Деление Arthroidignatha на Homopterida и Heteroptera. В этой классификации все Arthroidignatha, не относящиеся к голофилетическому таксону Heteroptera, объединяются в парафилетический таксон Homopterida Pearce 1936 {по циркумскрипции соответствует «Homoptera» sensu Westwood 1838 (nec Latreille 1810)}. Этот таксон характеризуется следующими плезиоморфиями, отличающими его от Heteroptera.
(1) Внешний хоботок [см. Arthroidignatha (1)] отходит от задней части головы, позади него нет гулы. Очевидно, это плезиоморфия, поскольку внешний хоботок образован нижней губой, которая исходно является слитыми конечностями последнего сегмента головы и своим основанием граничит непосредственно с шейной мембраной.
(2) Передние крылья не разделены на кориум и перепоночку (как у всех прочих насекомых кроме Heteroptera).
II. Деление Arthroidignatha на Hemelytrata и Plantisuga. В то время как таксон Homopterida характеризуется лишь плезиоморфиями, таксоны Hemelytrata и Plantisuga имеют апоморфии, свидетельствующие об их голофилии (см. ниже).
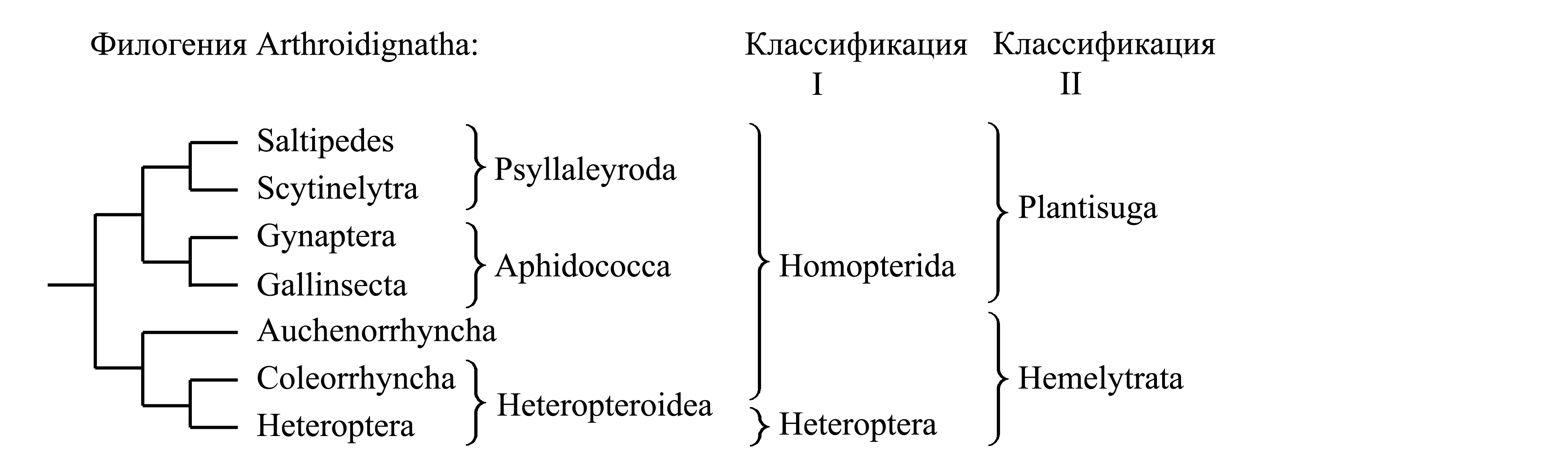
Здесь мы принимаем деление савременных Arthroidignatha на два подчиненных таксона – Hemelytrata и Plantisuga (стр. 000).
!!!!!ископаемые Protopsyllidioidea###???
Arthroidignatha, или Cimex/f2=Cicada/g4
![]() 1.
Hemelytrata,
или Cimex/f3=Cicada/g5
1.
Hemelytrata,
или Cimex/f3=Cicada/g5
![]() 1.1. Auchenorrhyncha, или
Cicada/f1=g6
1.1. Auchenorrhyncha, или
Cicada/f1=g6
![]() 1.1.1. Subtericornes, или
Fulgora/fg1
1.1.1. Subtericornes, или
Fulgora/fg1
![]() 1.1.2. Euhomoptera, или Cicada/f2=g7
1.1.2. Euhomoptera, или Cicada/f2=g7
![]() 1.2. Heteropteroidea, или
Cimex/f4=Notonecta/g1
1.2. Heteropteroidea, или
Cimex/f4=Notonecta/g1
![]() 1.2.1. Coleorrhyncha, или
Peloridium/fg1
1.2.1. Coleorrhyncha, или
Peloridium/fg1
![]() 1.2.2. Heteroptera, или Cimex/f5=Notonecta/g2
1.2.2. Heteroptera, или Cimex/f5=Notonecta/g2
![]() 1.2.3. †Progonocimex/fg1
1.2.3. †Progonocimex/fg1
![]() 2. Plantisuga, или Aphis/fg1
2. Plantisuga, или Aphis/fg1
![]() 2.1. Psyllaleyroda, или Psylla/fg1
2.1. Psyllaleyroda, или Psylla/fg1
![]() 2.1.1. Saltipedes, или Psylla/fg2
2.1.1. Saltipedes, или Psylla/fg2
![]() 2.1.2. Scytinelytra, или
Aleyrodes/fg1
2.1.2. Scytinelytra, или
Aleyrodes/fg1
![]() 2.2. Aphidococca, или
Aphis/fg2
2.2. Aphidococca, или
Aphis/fg2
![]() 2.2.1. Gynaptera, или Aphis/fg3
2.2.1. Gynaptera, или Aphis/fg3
![]() 2.2.2. Gallinsecta, или Coccus/fg1
2.2.2. Gallinsecta, или Coccus/fg1