КЛАДОЭНДЕЗИС НАСЕКОМЫХ

![]()
|
КЛАДОЭНДЕЗИС НАСЕКОМЫХ |
|||||||
|
|
|
fg | |||||
IX-1.2. Acercaria Börner 1904
(Somobialia![]() Eucaryota
Eucaryota![]() Metazoa
Metazoa![]() Monostomata
Monostomata![]() Haemataria
Haemataria![]() Peritoneata
Peritoneata![]() Metameria
Metameria![]()
Gnathopoda![]() Euarthropoda
Euarthropoda![]() Mandibulata
Mandibulata![]() Atelocerata
Atelocerata![]() Hexapoda
Hexapoda![]() Amyocerata
Amyocerata![]()
Pterygota![]() Metapterygota
Metapterygota![]() Neoptera
Neoptera![]() Eumetabola
Eumetabola![]() Parametabola
Parametabola![]() Acercaria)
Acercaria)
(рис. 9.2.1–9.12.4)
|
Названия, однозначно соответствующие по циркумскрипции: — Acercaria Börner 1904a; — Anurota Chen 1962; — Acercarida Boudreaux 1979b; — Hemineoptera Kukalova-Peck & Brauckmann 1992. Названия, неоднозначно соответствующие по циркумскрипции (соответствующие также Parametabola): — Panhomoptera Crampton 1918, s.str. — Paraneoptera Martynov 1923, s.str. Иерархическое название: Pediculus/f10=Cicada/g2 [f:1777; g:L.1758(gen.195)] (sine Zorotypus). |
|
Ранг. От надотряда до надкогорты.
Название. Название Acercaria следует считать соответствующим по циркумскрипции данному таксону, несмотря на то, что исходно в состав Acercaria был включен таксон Conorrhyncha Börner 1904a {nom.typ.: Thaumatoxena/fg [fg:1904a]}. Единственный вид, относимый к Conorrhyncha – Thaumatoxena wasmanni Börner 1904, был тогда только что открыт и недостаточно изучен; Бернер (Börner 1904a) исходно отнес его к отряду «Rhynchota» в составе Condylognatha в составе Acercaria в составе Hemimetabola, что следует считать фактической ошибкой, а не мнением о классификации. В следующей работе (Börner 1908) он исправил ошибку и перенес Thaumatoxena/fg в отряд Diptera.
Аутапоморфии Acercaria. Отдельные части ротового аппарата видоизменены: максиллы имеют уникальное строение [см. (1)], нижняя губа упрощена [см. (3)], клипеус и цибариальные мышцы увеличены [см. (4)]. К Acercaria относятся как насекомые с сосущим ротовым аппаратом (трипсы и членистохоботные), у которых все эти особенности строения ротовых частей имеют большое функциональное значение [см. ниже, Condylognatha (1)], так и насекомые, сохраняющие грызущий ротовой аппарат – сеноеды и пухоеды (Parapsocida, Eupsocida и Mallophaga). Отсюда можно заключить, что описанные ниже преобразования ротовых частей исходно не были связаны с сосущей специализацией и, по-видимому, исходно вообще не имели функционального значения.
(1) Модификация максилл. Максиллы имеют уникальное строение, с втягивающейся и выдвигающейся лацинией [не имеющей зубцов и щетинок на медиальной стороне – см. Parametabola (1)].
(1 a) Особенности строения максиллы, общие для Acercaria. Максиллярная лациния сильно удлинена, утончена и склеротизована в виде штыря (у Parapsocida и Eupsocida) или стилета (у Condylognatha), основание которого погружено глубоко в голову. Проксимальная погруженная часть лацинии заключена в трубчатый футляр, образованный мембранозной кутикулой, соединяющей основание лацинии с внешним покровом. Благодаря эластичности лациниального футляра лациния способна вдвигаться и выдвигаться. Мышца, идущая от основания лацинии в стипес, работает как протрактор лацинии, а мышца, идущая от основания лацинии к краниуму, работает как ретрактор лацинии (рис. 9.4.1B–C). Мышцы, двигающие лацинию, гомологичны стипито-лациниальной и кранио-лациниальной мышцам других насекомых (рис. 9.4.1A) [см. Hexapoda (2a)], но из-за изменения формы и прикрепления лацинии они принципиально изменили свою функцию: вместо того, чтобы сдвигать лацинии навстречу друг другу, они работают к антагонисты, вдвигающие и выдвигающие лацинию.
Помимо модификации лацинии, особенностью максилл всех Acercaria является утрата кардо, так что стипес либо непосредственно сочленяется с головной капсулой, либо слит с головной капсулой неподвижно.
Среди Acercaria максиллы с их выдвигающимися лациниями по-разному функционируют в разных таксонах: у сеноедов лациния способствуют движениям дистальной доли максиллы – малы [см. ниже, (1b)]; у Parasita лацинии не функционируют, вестигиальные или совсем утрачены; у трипсов и членистохоботных лацинии левой и правой максилл, складываясь вместе, образуют выдвигающуюся сосательную трубку, работающую в составе колюще-сосущего ротового аппарата [см. ниже, Condylognatha (1b)].
(1 b) Исходное строение максиллы. Судя по всему, исходное для Acercaria строение максиллы имеется у Panpsocoptera, поскольку их ротовой аппарат сохраняет исходную для насекомых грызущую функцию и включает нормально развитые подвижные мандибулы с двухмыщелковым причленением и выраженными инцизорами и молами [см. Metapterygota (2) и Mandibulata (1)]. При этом полный набор мышц, исходно присущий максилле насекомых [см. Hexapoda (2)], сохраняется у примитивных сеноедов – Parapsocida (рис. 9.2.4A), тогда как у настоящих сеноедов (Eupsocida) не хватает стипито-галеальной мышцы (рис. 9.2.4B), а у пухоедов не хватает лациниальных мышц (рис. 9.3.1B).
У Parapsocida (рис. 9.2.4A) стипес максиллы расчленен на две доли – проксимальную – базистипес (basistipes) и дистальную – максиллярную малу (mala maxillaris); при этом расчленении место прикрепления щупика оказалось на базистипесе, а место прикрепления лацинии находится на мале; галеа не выражена. Основания всех мышц, исходно идущих от стипеса к тенториуму, крепятся в базистипесе. Основания мышц, идущих от стипеса к щупику, также крепятся в базистипесе. К втягивающейся лацинии подходят 3 мышцы: кранио-лациниальная мышца (служащая ретрактором) (рис. 9.2.4A: cr-lac) и две мышцы, соответствующие одной исходной стипито-лациниальной мышце – одна из них крепится в базистипесе, другая в мале (рис. 9.2.4A: stp-lac1, stp-lac2). От базистипеса к основанию малы идет мышца, соответствующая исходной стипито-галеальной мышце насекомых (рис. 9.2.4A: stp-mal); из этого следует, что галеа вошла в состав малы. Малу иногда считают галеей на том основании, что она двигается стипито-галеальной мышцей; однако истинная галеа не может служить местом прикрепления лацинии и лациниальной мышцы. Судя по всему, подвижное сочленение между базистипесом и малой возникло у общего предка Acercaria и не соответствует исходному для насекомых месту сочленения между стипесом и галеей.
Мала соединена с базистипесом подвижным одномыщелковым сочленением, расположена почти под прямым углом к базистипесу и направлена вершиной медиально, так что вершины левой и правой мал могут смыкаться и прижиматься друг к другу. Вершина малы бывает пузыревидной, мембранозной (рис. 9.2.4A–B), либо несет зубчики, изогнутые проксимально (рис. 9.3.1B). С помощью такой малы максилла способна совершать загребательные движения и перемещать в рот частицы пищи, разгрызенной мандибулами. Движения малы могут обеспечиваться лишь двумя мышцами, которые крепятся к ее основанию – это мышца, идущая из базистипеса к мале (исходная стипито-галеальная мышца) и мышца, идущая от малы к проксимальному концу втягивающейся лацинии (часть исходной стипито-лациниальной мышцы); обе мышцы могут служить флексорами малы, т.е. поворачивать малу ко рту; обратное движение малы возможно только гидравлическим способом.
Для того, чтобы использовать для сгибания малы мышцу, идущую от нее к лацинии, сеноед должен зафиксировать лацинию неподвижно. Это можно сделать, если с помощью мышцы, идущей от лацинии к базистипесу, выдвинуть лацинию (как в правой половине рис. 9.2.4B) и упереться ее вершиной в субстрат. О том, что такой механизм используется сеноедами, косвенно свидетельствует тот факт, что у всех видов сеноедов (как Eupsocida, так и Parapsocida) выдвигающиеся лацинии максилл одинаково хорошо развиты, штыревидной формы и имеют примерно одинаковую длину относительно максилл; в отличие от Parasita, они никогда не подвергаются редукции; вершины лациний у разных сеноедов видов могут различаться по форме, но всегда имеют острия, направленные дистально; это позволяет упирать их в субстрат и использовать как опору для мышцы, идущей от лацинии к мале. Высказывалось предположение, что штыревидные лацинии сеноедов служат как долота для дробления твердой пищи; однако у всех сеноедов мышцы, выдвигающие и втягивающие лацинии, во много раз тоньше, чем мышцы, сжимающие и раздвигающие мандибулы (ср. рис. 9.2.4B и D); так что любую пищу гораздо легче измельчить мандибулами, чем лациниями максилл.
Способность максиллы совершать сложные движения, загребающие пищу в рот, необходима для насекомого с грызущими мандибулами, поскольку мандибула [одночлениковая и имеющая два мыщелка – см. Mandibulata (1b)] к таким движениям неспособна. Исходно у насекомых максилла имеет два подвижно сочлененных склерита – кардо и стипес, что позволяет максиллам совершать необходимые загребающие движения [см. Hexapoda (2a)] (рис. 9.1.3F). У Acercaria расчленение максиллы на кардо и стипес утрачено, но это компенсируется ее расчленением на базистипес и малу. В отличие от кардо и стипеса, к каждому из которых исходно подходят мышцы от эндоскелета головы, к мале такие мышцы не подходят. Поэтому мала должна иметь способность двигаться либо с помощью мышцы, идущей к ней от базистипеса, либо с помощью сложного механизма, включающего выдвигающуюся лацинию. У Parapsocida имеются оба эти механизма – и мышца, идущая из базистипеса, и лациния с полным набором мышц.
(1 с) Модификации в строении максиллы среди Acercaria. В отличие от Parapsocida, имеющих полный набор максиллярных мышц (рис. 9.2.4A), у настоящих сеноедов (Eupsocida) утрачена мышца, идущая от базистипеса к мале, так что движение малы может осуществляться только с помощью выдвигающейся лацинии [см. ниже, Eupsocida (2) и рис. 9.2.4B–C]. У пухоедов, наоборот, лациния подверглась редукции, и мала двигается только мышцей, идущей к ней от базистипеса [см. ниже, Parasita (5) и рис. 9.3.1B]. У трипсов и членистохоботных ротовой аппарат преобразовался в колюще-сосущий, мандибулы утратили жевательную функцию, и необходимость в подвижных малах исчезла; малы утратились, а лацинии преобразовались в выдвигающийся сосущий хоботок [см. ниже, Condylognatha (1b), Arthroidignatha (1b) и рис. 9.4.1С]. У некоторых членистохоботных с десклеротизованным краниумом основания краниально-лациниальных мышц частично или полностью переместились с краниума на тенториум [см. ниже, Plantisuga (1)].
Несмотря на то, что полость максиллярной лацинии сильно сужена, у сеноедов сохраняется нормальная линька, при которой новая лациния закладывается внутри кутикулы лацинии предыдущего возраста; у членистохоботных, имеющих особенно узкую полость лацинии, линька модифицирована и новая лациния закладывается проксимальнее старой, закручиваясь в полости головы плоской спиралью (Weber 1929).
(2) Четырехчлениковый максиллярный щупик. Максиллярный щупик 4-члениковый или с меньшим числом члеников [в отличие от пятичленикового у Zoraptera и многих других – см. Hexapoda (2c)].
4-члениковые щупики имеются у Parapsocida, Eupsocida и части Amblycera (рис. 9.2.4, 9.3.1B); у некоторых Amblycera число члеников уменьшено до 3 или 2; у прочих Parasita максиллярные щупики утрачены. У Thysanoptera щупик с 3 или меньшим числом первичных члеников; иногда имеется вторичное расчленение, так что щупик кажется имеющим до 8 члеников [см. ниже, Thysanoptera (2)]. У Arthroidignatha щупики утрачены.
(3) Модификация лабиума. Нижняя губа [см. Hexapoda (3)] упрощенного строения: глоссы срослись в непарную алаглоссу; щупики [исходно трехчлениковые – см. Hexapoda (3c)] 2-члениковые. В некоторых таксонах Acercaria нижняя губа претерпела более глубокие модификации, при которых алаглосса и параглоссы бывают утрачены, а щупики не расчленены или утрачены.
Среди Panpsocoptera 2-члениковые лабиальные щупики имеются у части Parapsocida, Eupsocida и Amblycera; у части Parapsocida, Eupsocida, Amblycera и всех Ischnocera лабиальные щупики одночлениковые (рис. 9.3.1A); у слоновьих вшей щупики вестигиальные, у вшей щупики утрачены [см. ниже, Rhyncophthirina (1) и Siphunculata (1f)]. У трипсов щупики бывают вторично расчленены, так что лабиальные щупики бывают 1–5-члениковые [см. ниже, Thysanoptera (2)] (рис. 9.5.2). У членистохоботных вся нижняя губа преобразована в непарный внешний хоботок [см. ниже, Arthroidignatha (1d)].
(4) Увеличение клипеуса. Наличник [служащий местом прикрепления цибариальных мышц и исходно располагающийся вентральнее передних тенториальных ямок – см. Amyocerata (3)] увеличен и занимает большую часть лица, медиально заходит дорсальнее передних тенториальных ямок и вдается между основаниями антенн. Клипео-цибариальные мышцы соответствующим образом увеличены и образуют конический пучок, широкое основание которого крепится к клипеусу, а узкая вершина к непарному склериту на передней (дорсальной) стенке цибариума. У представителей с сосущим ротовым аппаратом мощные клипео-цибариальные мышцы используются для сосания жидкой пищи; однако исходно увеличение этих мышц, судя по всему, не связано с сосущей специализацией, поскольку свойственно также сеноедам и пухоедам (Parapsocida, Eupsocida и Mallophaga), сохраняющим грызущий ротовой аппарат. Наоборот, у некоторых представителей с сосущим ротовым аппаратом клипеус вторично уменьшен [в частности, см. ниже, Subtericornes (1)].
Среди Acercaria наименее специализированный ротовой аппарат имеется у сеноедов (Parapsocida и Eupsocida). У них клипеус обращен вперед, сильно выпуклый, полушаровидный; на передней стенке глотки, куда сходятся мощные клипео-цибариальные мышцы, имеется непарный пальцевидный склеротизованный выступ (рис. 9.2.3). Некоторые авторы ошибочно считают, что этот пальцевидный склерит входит в углубление на ситофорном склерите противоположной стенки глотки [см. ниже, Panpsocoptera (1)] и способен дробить твердые частицы пищи (Weber 1933, Snodgrass 1944). Однако на самом деле мышцы не могут толкать; стенки глотки мембранозные и также не могут вталкивать пальцевидный склерит в углубление ситофорного склерита. При плотно сдвинутых мандибулах их молы смыкаются в полости глотки, разделяя пальцевидный и ситофорный склериты (рис. 9.2.3).
У Mallophaga-Ischnocera клипеус также обращен вперед, выпуклый, а отходящие от него клипео-цибариальные мышцы конически сходятся на пальцевидном цибариальном склерите, напротив которого находится углубление ситофорного склерита (рис. 9.3.1D).
У Mallophaga-Amblycera (также имеющих грызущий ротовой аппарат) голова прогнатная, клипеус смещен на дорсальную сторону, уплощен и не отделен ото лба; область прикрепления клипео-цибариальных мышц к глотке вытянута вдоль глотки, а цибариальный выступ дорсальной стенки глотки утратил пальцевидную форму и имеет вид продольного гребня (рис. 9.3.1C).
У Siphunculata, Rhyncophthirina и Condylognatha, имеющих независимо возникшие сосущие ротовые аппараты, выпуклый склерит на дорсальной (передней) стенки глотки, оттягиваемый мощными клипео-цибариальными мышцами, выполняет функцию глоточного насоса. У вшей и слоновьих вшей (Siphunculata и Rhyncophthirina), как и у Amblycera, голова прогнатная, увеличенный клипеус находится на ее дорсальной стороне, уплощен и не отделен ото лба.
У различных Condylognatha клипеус бывает отделен или не отделен ото лба, и занимает большую или меньшую площадь на вентральной, передней и/или дорсальной стороне головы.
(5) Утрата церков. Церки [см. Metapterygota (4)] полностью утрачены (с чем связано название). Неуникальная апоморфия (см. Указатель признаков [3.41]).
(6) Уменьшение числа мальпигиевых сосудов. Число мальпигиевых сосудов [исходно стабильное и уменьшенное – см. Eumetabola (5) и Parametabola (7)] сокращено до 4 штук (т.е. до 2 пар): у всех Panpsocoptera, Thysanoptera, Hemelytrata и Saltipedes имеется 4 (т.е. 2 пары) мальпигиевых сосудов; у Scytinelytra и большинства Gallinsecta число мальпигиевых сосудов сокращено до 2 (т.е. 1 пары); у Gynaptera мальпигиевы сосуды утрачены. Те же числа мальпигиевых сосудов встречаются в некоторых других таксонах (см. Указатель признаков [4.7]: табл. F).
(7) Мицетомы. В брюшке имеются мицетомы – тела, в которых находятся симбиотические бактерии (название «мицетомы» связано с существовавшим ранее мнением, что часть симбионтов относится к бактериям, а часть к дрожжевым грибкам). Симбионты находятся преимущественно внутри клеток – мицетоцитов, большей частью преобразованных в синцитии. Мицетомы наследуются, передаваясь от самки к яйцу при оогенезе. В одном насекомом может существовать несколько видов симбионтов, различающихся по местоположению в организме и способу передачи в яйцо.
У сеноедов (Parapsocida и Eupsocida) мицетомы не обнаружены; мицетомы единой природы обнаружены у Parasita и у многих Arthroidignatha – у всех исследованных в этом отношении Gynaptera, Scytinelytra, Saltipedes, Gallinsecta, Auchenorrhyncha, а так же у Coleorrhyncha и некоторых Heteroptera (Емельянов 1987).
Геологический возраст. Пермь – ныне (Rasnitsyn & Quicke 2002).
Систематическое положение Acercaria. Здесь мы рассматриваем Acercaria как подчиненный таксон в составе Parametabola. Некоторые авторы не признают таксон Parametabola, считая, что Zoraptera не имеют родства с Acercaria (см. Zoraptera: «Систематическое положение» II–V). В соответствии с этим вместо таксона Eumetabola, объединяющего Metabola с Parametabola, выделяют таксон Phalloneopterata, объединяющий Metabola с Acercaria (см. Eumetabola: «Статус»).
I. Деление Acercaria на Micracercaria и Arthroidignatha. Таксон Micracercaria Willmann 2003 объединяет Panpsocoptera и Thysanoptera. В качестве их синапоморфий было указано только сходство в конфигурации мышцы цибариального насоса и наличие двух аксонем в жгутике сперматозоида. В действительности цибариальные мышцы Copeognatha и Thysanoptera не имеют общих особенностей, отсутствующих у Arthroidignatha [см. Acercaria (4)]. Строение сперматозоидов, будучи изученным для небольшого числа видов, имеет большое видовое разнообразие; представители с увеличенным числом аксонем встречаются в неродственных друг другу таксонах насекомых (Dollai et al. 2016). Выделение таксона Micracercaria противоречит представлению о голофилии таксона Condylognatha.
II. Деление Acercaria на Panpsocoptera и Condylognatha. Эта классификация основана на уникальных синапоморфиях в строении ротовых аппаратов трипсов и членистохоботных [см. ниже, Condylognatha (1)].
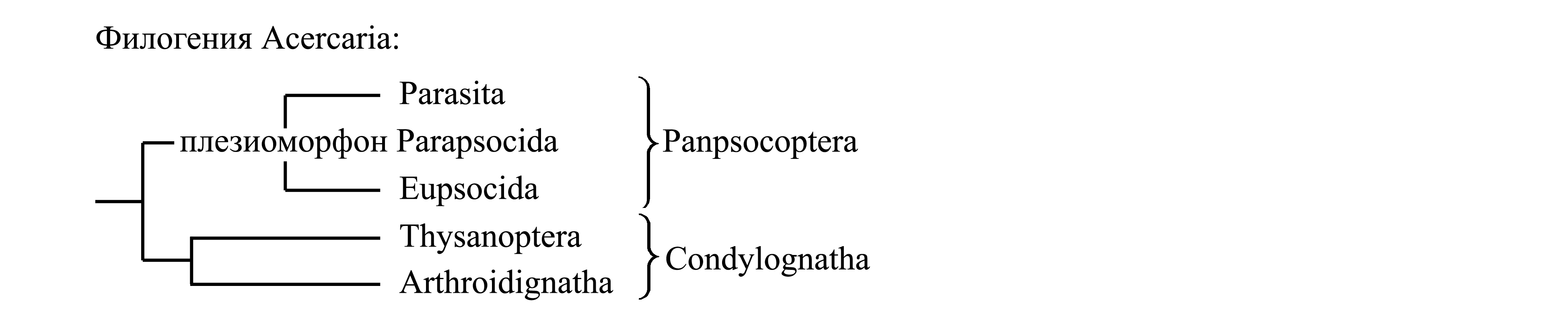
Здесь мы принимаем деление Acercaria на два подчиненных таксона – Panpsocoptera и Condylognatha (стр. 0000).
Acercaria, или Pediculus/f10=Cicada/g2
![]() 1. Panpsocoptera, или Pediculus/f11=g1
1. Panpsocoptera, или Pediculus/f11=g1
![]() 1.1. Плезиоморфон Parapsocida, или
Atropos/f1=Trogium/g1
1.1. Плезиоморфон Parapsocida, или
Atropos/f1=Trogium/g1
![]() 1.2. Eupsocida, или Psocus/fg1
1.2. Eupsocida, или Psocus/fg1
![]() 1.3. Parasita, или Pediculus/f12=g2
1.3. Parasita, или Pediculus/f12=g2
![]() 1.3.1. Mallophaga, или
Philopterus/f1=Ricinus/g1
1.3.1. Mallophaga, или
Philopterus/f1=Ricinus/g1
![]() 1.3.1.1. Amblycera, или Ricinus/f1=g2
1.3.1.1. Amblycera, или Ricinus/f1=g2
![]() 1.3.1.2. Ischnocera, или
Philopterus/f2=g1
1.3.1.2. Ischnocera, или
Philopterus/f2=g1
![]() 1.3.2. Siphunculata, или
Pediculus/f13=g3
1.3.2. Siphunculata, или
Pediculus/f13=g3
![]() 1.3.3. Rhyncophthirina, или
Haematomyzus/fg(1)
1.3.3. Rhyncophthirina, или
Haematomyzus/fg(1)
![]() 2. Condylognatha, или Cimex/f1=Cicada/g3
2. Condylognatha, или Cimex/f1=Cicada/g3
![]() 2.1. Thysanoptera, или Thrips/fg1–2
2.1. Thysanoptera, или Thrips/fg1–2
![]() 2.2. Arthroidignatha, или
Cimex/f2=Cicada/g4
2.2. Arthroidignatha, или
Cimex/f2=Cicada/g4
![]() 2.2.1. Hemelytrata, или Cimex/f3=Cicada/g5
2.2.1. Hemelytrata, или Cimex/f3=Cicada/g5
![]() 2.2.1.1. Auchenorrhyncha, или
Cicada/f1=g6
2.2.1.1. Auchenorrhyncha, или
Cicada/f1=g6
![]() 2.2.1.1.1. Subtericornes, или
Fulgora/fg1
2.2.1.1.1. Subtericornes, или
Fulgora/fg1
![]() 2.2.1.1.2. Euhomoptera, или
Cicada/f2=g7
2.2.1.1.2. Euhomoptera, или
Cicada/f2=g7
![]() 2.2.1.2. Heteropteroidea, или
Cimex/f4=Notonecta/g1
2.2.1.2. Heteropteroidea, или
Cimex/f4=Notonecta/g1
![]() 2.2.1.2.1. Coleorrhyncha, или
Peloridium/fg1
2.2.1.2.1. Coleorrhyncha, или
Peloridium/fg1
![]() 2.2.1.2.2. Heteroptera, или
Cimex/f5=Notonecta/g2
2.2.1.2.2. Heteroptera, или
Cimex/f5=Notonecta/g2
![]() 2.2.1.2.3. †Progonocimex/fg1
2.2.1.2.3. †Progonocimex/fg1
![]() 2.2.2. Plantisuga, или Aphis/fg1
2.2.2. Plantisuga, или Aphis/fg1
![]() 2.2.2.1. Psyllaleyroda, или
Psylla/fg1
2.2.2.1. Psyllaleyroda, или
Psylla/fg1
![]() 2.2.2.1.1. Saltipedes, или
Psylla/fg2
2.2.2.1.1. Saltipedes, или
Psylla/fg2
![]() 2.2.2.1.2. Scytinelytra, или
Aleyrodes/fg1
2.2.2.1.2. Scytinelytra, или
Aleyrodes/fg1
![]() 2.2.2.2. Aphidococca,
или Aphis/fg2
2.2.2.2. Aphidococca,
или Aphis/fg2
![]() 2.2.2.2.1. Gynaptera, или
Aphis/fg3
2.2.2.2.1. Gynaptera, или
Aphis/fg3
![]() 2.2.2.2.2. Gallinsecta, или
Coccus/fg1
2.2.2.2.2. Gallinsecta, или
Coccus/fg1