КЛАДОЭНДЕЗИС НАСЕКОМЫХ
![]()
![]()
|
КЛАДОЭНДЕЗИС НАСЕКОМЫХ |
|||||||
|
|
|
fg | |||||
VIII-1.2.1. Plecoptera Burmeister 1839 – веснянки
(Somobialia![]() Eucaryota
Eucaryota![]() Metazoa
Metazoa![]() Monostomata
Monostomata![]() Haemataria
Haemataria![]() Peritoneata
Peritoneata![]() Metameria
Metameria![]()
Gnathopoda![]() Euarthropoda
Euarthropoda![]() Mandibulata
Mandibulata![]() Atelocerata
Atelocerata![]() Hexapoda
Hexapoda![]() Amyocerata
Amyocerata![]()
Pterygota![]() Metapterygota
Metapterygota![]() Neoptera
Neoptera![]() Rhipineoptera
Rhipineoptera![]() Plecoptera)
Plecoptera)
(рис. 8.3.1–8.3.3)
(рис. 8.3.1,
8.3.2, 8.3.3)
|
Названия, однозначно соответствующие по циркумскрипции: — Plecoptera Burmeister 1839; — Perloptera Krausse & Wolff 1919; — Plecopterida Pearce 1936; — Pleconeoptera Kukalova-Peck & Peck 1993. По циркумскрипции соответствует таксонам под родовым названием Perla (sensu Geoffroy 1764) и производными типифицированными названиями: Perlariae, Perlarinia, Perlarides, Perlides, Perlites, Perlina, Perlidae, Perlinae, Perlidi, Perloidea, Perlaria, Perlida. Иерархическое название: Perla/fg1 [f:1802; g:1762] (incl. Nemoura). |
|
Ранг. В XX веке – отряд или надотряд; в XIX веке – обычно от семейства до отряда; в XVIII веке – род.
Аутапоморфии Plecoptera
(1) Метаморфоз. Личинки и имаго адаптированы к обитанию в разных средах: имаго сохраняют наземный образ жизни и дышат воздухом, а личинки обитают в пресных водоемах и дышат растворенным в воде кислородом. Помимо веснянок, подобный амфибиотический образ жизни свойственен только поденкам (Ephemeroptera), стрекозам (Odonata) и некоторым насекомым с полным превращениям (Metabola); у них обитание личинок и имаго в разных средах связано с глубокими морфологическими различиями между личинками и имаго, во многих случаях с потерей эволюционной корреляции между личиночными и имагинальными признаками [ср.: Ephemeroptera (1)–(13), Odonata (1)–(10) и Metabola (1)]. В отличие от них, у веснянок морфологические различия между личинками и имаго минимальны (рис. 8.3.1 и 8.3.2A): помимо общих для Pterygota различий в строении груди [см. Pterygota (4c)] и отсутствия у личинки развитых гениталий, личинки и имаго веснянок различаются строением претарсуса [см. (1A)] и абдоминальных мышц [см. (1B)]. Другие различия, если имеются, присущи лишь отдельным таксонам веснянок и не являются исходными для Plecoptera. Несмотря на обитание в разных средах, личинки и имаго изначально имеют одинаковый ротовой аппарат и питаются одинаковой пищей; лишь в отдельных таксонах возникло различие между личинками и имаго в этом отношении [см. (1C)].
Личинки большинства видов приспособлены к обитанию в быстрых водотоках с высоким содержанием кислорода (преимущественно горных и предгорных ручьях и реках), и лишь немногие виды обитают в медленных равнинных реках. В связи с подводным образом жизни у личинок трахеостии в значительной части нефункционирующие [но при каждой линьке старая кутикула трахей выходит через все 10 пар трахеостий – см. Hexapoda (9b)]; тем не менее грудные и передние пары брюшных трахеостий могут функционировать как дыхальца. Благодаря этому для линьки на имаго личинка способна выйти из воды; иногда личинка проходит значительное расстояние по суше, прежде чем полинять на имаго. У некоторых Gripopteryx/f=Leptoperla/g личинки вторично перешли к наземному образу жизни. Личинки некоторых видов имеют жабры в виде тонкостенных выростов на тех или иных частях тела [см. (1D)].
В связи с амфибиотическим образом жизни и обитанием личинок в быстрых потоках у некоторых веснянок имеется необычный половой диморфизм в развитии крыльев у имаго [см. (1E)].
Число личиночных возрастов большое и неопределенное, от 10 до 22 (Stewart & Stark 1993).
(1 A) Претарсус личинок и имаго. Претарсус несет два коготка [см. Pterygota (5)], форма которых одинакова у личинки и у имаго. У имаго на вентральной сочленовной мембране каждого коготка имеется крупная щетинка (называемая базипульвиллой), а между коготками находится пузыревидный аролий. У личинки нет ни аролия, ни базипульвилл. Помимо веснянок, аролий отсутствует у личинки и появляется у имаго у примитивных представителей Dermatoptera; у многих других насекомых личинки и имаго не различаются по этому признаку большинства других насекомых аролий одинаково развит у личинок и имаго [см. Pterygota (5)].
(1 B) Локомоция личинок. Личинки веснянок ведут донный образ жизни и по большей части передвигается с помощью ног. Но если личинка веснянки оказывается вынужденной плыть, она всегда изгибается в латеральном направлении, двигая брюшком из стороны в сторону. Поскольку личинки большинства веснянок уплощены дорсовентрально, такие движения оказываются наименее эффективными для плавания. Сходные движения присущи личинкам равнокрылых стрекоз; в отличие от веснянок, у этих стрекоз латеральные движения оказываются эффективными благодаря наличию плоских хвостовых жабр [ср.: Zygoptera (1)]. По способу плавания личинки веснянок резко отличаются от личинок поденок, для которых характерны дорсовентральные плавательные движения брюшком [ср.: Ephemeroptera (1e)].
У некоторых личинок веснянок имеются ряды или поля длинных, густо расположенных щетинок на туловище, ногах и/или церках. Эти щетинки не делают плавательные движения личинки более эффективными, но могут иметь иное гидродинамическое значение, т.к. способствуют прижиманию личинки к субстрату в быстром потоке. У имаго тех же видов этих щетинок нет. Такие щетинки, свойственные только стадии личинки, присущи лишь отдельным таксонам веснянок и не входят в общий план строения Plecoptera. В немногих таксонах веснянок личинки приобрели особую форму тела, отличную от имагинальной.
(1 C) Питание и ротовой аппарат у личинок и имаго. Из учебника в учебник кочует ошибочное утверждение, что отряд Plecoptera характеризуются афагией и редукцией ротового аппарата у имаго. В действительности у большинства веснянок (у Euholognatha и Antarctoplecoptera) ротовой аппарат личинок и имаго одинакового примитивного строения и используется для питания детритом и водорослями. Несмотря на то, что у многих веснянок голова прогнатная, гула всегда отсутствует (т.е. нет дополнительной склеротизации позади основания нижней губы – см. Указатель признаков [1.3]) (рис. 8.3.1). Ротовые части всегда обычного грызущего типа [см. Hexapoda (1a) и Rhipineoptera (3)]: имеются верхняя губа, грызущие мандибулы, максиллы с галеей, лацинией и 5-члениковым (редко 4-члениковым) щупиком, гипофаринкс и нижняя губа с глоссами, параглоссами и 3-члениковыми щупиками [см. Hexapoda (2) и (3)]. Лишь у Systellognatha личиночные ротовые части модифицированы для хищничества, а имагинальные не функционируют и могут быть уменьшены; но и в этом случае они сохраняются; лишь у Systellognatha-Subulipalpia редуцированы глоссы нижней губы, но сохраняются остальные части ротового аппарата.
(1 D) Жабры. Личинки многих видов имеют жабры – мягкие кустистые, либо нитевидные выросты на различных участках тела – на нижней губе, шее или груди [см. (1Da)], на ногах [см. (1Db)], по бокам брюшка [см. (1Dc)] или на вершине брюшка [см. (1Dd)]. Жабры, расположенные на разных частях тела, явно не гомологичны друг другу. У многих веснянок никаких жабр нет вообще. В том числе жабры не обнаружены и у древних ископаемых палеозойских и мезозойских веснянок. По всей видимости, жабры возникали независимо в отдельных таксонах веснянок, и их наличие не входит в исходный план строения Plecoptera. Парадоксальным является то, что личинки веснянок, дышащие только растворенным в воде кислородом, и в то же время имеющие плотные малопроницаемые покровы и зачастую крупные размеры, по большей части обходятся без специальных органов водного дыхания. Несмотря на очень долгую эволюцию (с ранней перми), те отдельные таксоны веснянок, которые приобрели жабры, не смогли вытеснить безжаберные формы.
Хотя все взрослые веснянки обитают на суше и имеют функционирующие дыхальца, у некоторых видов на стадии имаго сохраняются жабры, соответствующие личиночным и не резорбирующиеся при метаморфозе [ см. Hexapoda (9b)].
(1 D a) Вентральные жабры. У личинок разных таксонов в составе Euholognatha и Systellognatha развиты жабры на вентральной стороне головы и/или груди (Stewart & Stark 1993). В некоторых таксонах имеется пара нитевидных или кустистых жабр по бокам нижней губы или на шее. У некоторых Systellognatha на каждом из трех сегментов груди имеется по одной или по несколько пар нитевидных или кистевидных жабр; особенно хорошо развиты грудные жабры у Pteronarcys/fg и Perla/fg5 (sine Chloroperla, Perlodes). У перлид, помимо жабр на стернитах груди, бывает развита пара таких же жабр на первом стерните брюшка и/или пара кустистых жабр на парапроктах [см. (1Dd)]; у Pteronarcys/fg, помимо грудных жабр, имеются сходные по строению придыхальцевые жабры на I–III или I–II стернитах брюшка [см. (1Dc)].
(1 D b) Коксальные жабры. У личинок Taeniopteryx/fg (sine Brachypteryx) от медиальной стороны тазика каждой ноги отходит мембранозный отросток, состоящий из трех телескопически втягивающихся члеников (рис. 8.3.2A). Этот отросток, хотя и похож на ветвь конечности, на самом деле является не каким-то древним образованием, а жаброй уникального строения, возникшей лишь у данного частного таксона веснянок.
(1 D c) Парные брюшные жабры. Парные брюшные трахейные жабры, помимо личинок Pteronarcys/fg и Perla/fg5 [см. (1Da)], имеются у личинки североамериканского вида Oroperla barbara Needham 1933 (относящегося к Perlodes/fg) и у личинок амфинотического таксона Archiperlaria. У личинок Archiperlaria на боках первых четырех-шести сегментов брюшка имеется по паре кустистых или нитевидных жабр. Каждая такая жабра отходит от задне-вентро-латеральной части сегмента (позади и вентральнее трахеостии) и начинается толстым основанием; в нее входит мышца, идущая от тергита, благодаря чему жабры способны волнообразно колебаться из стороны в сторону (Zwick 1973, 2000).
Высказывалось предположение, что неподвижные жабры Pteronarcys/fg и подвижные жабры Archiperlaria гомологичны парным абдоминальным жабрам других насекомых и исходны для Pterygota. В действительности эти жабры являются придыхальцевыми трахейными жабрами, т.к. каждая жабра трахеируется от дыхальцевой трахеи того же сегмента. Подобные жабры, помимо личинок веснянок, независимо возникли у некоторых других насекомых [см. Hexapoda (9) и Metapterygota (6)]; по строению и мускулатуре они принципиально отличаются от тергалий личинок поденок [см. Metapterygota (6); ср.: Ephemeroptera (12)]. Большинство личинок веснянок не имеет парных брюшных жабр; у личинок вымерших веснянок, включая палеозойских, жабры также отсутствуют (Синиченкова 1987). Вероятно, парные брюшные жабры независимо возникли в разных таксонах веснянок.
(1 D d) Анальные жабры. У личинок большинства австроперлид (Austroperla/fg) на эпипрокте, на каждом парапрокте и на вершине каждого церка имеется мягкий членистый жаберный отросток, так что от конца брюшка назад отходят 5 членистых хвостовых придатков (рис. 8.3.3); у некоторых видов автроперлид, кроме того, имеется пара таких же отростков между эпипроктом и церками, так что всего имеется 7 членистых отростков. У относящегося к австроперлидам тасманийского вида Crypturoperla paradoxa Illies 1969 вместо членистых жаберных отростков имеется нечленистый нитевидный жаберный отросток на эпипрокте и пучки многочисленных нитевидных жаберных отростков на каждом парапрокте и на первом членике каждого церка (Illies 1969).
У личинок скопурид (Scopura/fg) на десятом уромере имеется 5 нечленистых нитевидных жаберных отростков: непарный отросток на эпипрокте, по одному отростку на основании каждого парапрокта и пара отростков латеральнее парапроктов (между парапроктами и тергитом); кроме того, на сочленовной мембране между девятым и десятым уромерами имеются многочисленные отростки такого же строения, скрывающиеся при втягивании десятого уромера в девятый.
У личинок грипоптеригид (Gripopteryx/f=Leptoperla/g) нет членистых жаберных отростков, но на вершине брюшка дорсальнее анального отверстия имеется пучок жаберных нитей, способный полностью или частично втягиваться внутрь брюшка под действием пары продольных дорсальных мышц. Эти мышцы начинаются от пары особых аподем, которые отходят от переднего края десятого тергита брюшка вперед и лежат под девятым тергитом брюшка; у некоторых видов эти аподемы укорочены, а мышцы, втягивающие жабры, начинаются частично от них, частично от девятого тергита брюшка (т.е. место прикрепления мышц частично сместилось с десятого уромера на девятый).
У личинок многих перлид [Perla/fg (sine Chloroperla, Perlodes)] на конце брюшка имеются кустистые жабры принципиально иного происхождения: они парные, расположены на парапроктах, и втягиваются короткими мышцами, проходящими внутри парапроктов и происходящими из продолжений вентральных продольных мышц, идущих от IX стернита брюшка к основаниям парапроктов [см. Amyocerata (8)].
По-видимому, все эти анальные жабры возникли независимо в разных таксонах веснянок. Высказывалось предположение, что непарный жаберный отросток, отходящий от эпипрокта у личинок австроперлид и скопурид, гомологичен парацерку Ephemeroptera и Triplura: как и парацерк, он находится дорсальнее анального отверстия, и к его основанию подходят мышцы, начинающиеся от десятого тергита брюшка. Однако в отличие от настоящего парацерка, непарный отросток эпипрокта у личинок веснянок всегда имеет строение отличное от строения церков и сходное со строением отростков парапроктов и других жаберных отростков того же вида. У личинок веснянок отросток эпипрокта может иметь эволюционную корреляцию с отростками парапроктов, но не с церками (рис. 8.3.3) [см. Metapterygota (4)].
(1 E) Половой диморфизм в развитии крыльев. У разных видов веснянок встречается такой половой диморфизм, при котором самцы короткокрылые или бескрылые, а самки имеют вполне развитые крылья. Вероятно, это связано с тем, что оплодотворенные самки должны для откладки яиц лететь вверх по течению, чтобы компенсировать постоянный снос личинок вниз по течению. У большинства других насекомых тенденцию к редукции крыльев имеют не самцы, а самки. У многих веснянок полового диморфизма в строении крыльев нет, и оба пола бывают полнокрылыми, короткокрылыми или бескрылыми.
(2) Трехчлениковые лапки. Лапки [исходно пятичлениковые – см. Amyocerata (5)] у имаго и личинок 3-члениковые (неуникальная апоморфия – Указатель признаков [2.50]). При этом неизвестно, какие из исходных пяти члеников слились или исчезли, поскольку ни у каких веснянок нет следов двух утраченных члеников. У личинки I возраста первые два членика очень короткие: вместе взятые они в несколько раз короче третьего членика. В различных таксонах у зрелых личинок и у имаго либо сохраняются те же пропорции, либо первый членик удлиняется; у имаго Taeniopteryx/fg все три членика одинаково длинные (рис. 8.3.1A). У некоторых видов первые два членика лапки, будучи короткими, слиты друг с другом, и в этом случае лапка 2-члениковая.
(3) Поперечные жилки преклавальной области крыла. На переднем крыле частые поперечные жилки обычно имеются в двух соседних полях – между продольными жилками M и Cu1 и между Cu1 и Cu2; в случаях, когда в других полях крыла поперечных жилок мало, эти два ряда поперечных жилок по обе стороны жилки Cu1 заметно выделяются, придавая переднему крылу веснянок характерный вид (рис. 8.3.1). На заднем крыле, в отличие от переднего, этих поперечных жилок нет. Исключение составляют Capnia/fg, у которых данные ряды поперечных жилок на переднем крыле редуцированы (так что между M и Cu1 имеется не более двух, а между Cu1 и Cu2 – не более одной поперечной жилки).
В отличие от поперечных, продольные жилки преклавальной области имеют обычное расположение. На переднем крыле RS отходит от RA, а на заднем крыле – от M [см. Rhipineoptera (1b)] (рис. 8.3.1, 8.3.2D–E); в остальном расположение преклавальных продольных жилок на передних и задних крыльях сходно. Жилка Sc впадает Y-образно и в C (передний край крыла), и в жилку RA. Жилка RA идет почти до вершины крыла. Жилка M (на переднем крыле) или общий ствол M+RS (на заднем крыле) начинается либо самостоятельно, либо отходит от жилки RA. Жилка Cu1 отходит Y-образно и от M, и от Cu2. В средней или дистальной части крыла продольные жилки RA, RS, M, Cu1 и Cu2 соединены поперечным кордом – ломаной линией из постоянных поперечных жилок; дистальнее поперечного корда жилки RS, M и Cu1 иногда дихотомически ветвятся, число их ветвей может быть изменчивым.
(4) Анальный веер. Анальный веер [имеющийся на заднем крыле – см. Rhipineoptera (1)] отличается по строению от анального веера Tegminoptera тем, что число его складок всегда равно трем и может не соответствовать числу веерообразно расходящихся жилок, причем передняя складка проходят не между жилками, а пересекает жилку V1 (рис. 8.3.1) [ср.: Tegminoptera (3)].
И на переднем, и на заднем крыле жилки Pcu и V1 [две передние жилки клавуса – см. Neoptera (1b)] около основания жестко соединены постоянной поперечной жилкой, так что имеется постоянная базальная анальная ячейка; в связи с этим на заднем крыле инверталярная складка [отделяющая анальный веер – см. Rhipineoptera (1)] в проксимальной части крыла огибает базальную анальную ячейку позади жилки V1, а далее пересекает жилку V1. Помимо инверталярной складки, обеспечивающей переворачивание всего анального веера [см. Neoptera (1b)], на анальном веере проходят еще две складки – вогнутая и выпуклая, благодаря чему анальный веер складывается втрое. На границе анального веера задний край крыла обычно образует характерную выемку (лишь у Archiperlaria этой выемки нет, и весь задний край с анальным веером равномерно-выпуклый).
В отличие от Tegminoptera, краевая жилка [являющаяся продолжением костальной жилки – см. Pterygota (1Ad)] проходит по всему краю крыла, как заднего, так и переднего, так что задний край анального веера также обрамлен тонкой краевой жилкой.
У некоторых веснянок (некоторые Capnia/fg и Chloroperla/fg) анальный веер подвергся редукции или совсем утрачен. Однако во всех случаях сохраняется в той или иной мере выраженное различие переднего и заднего крыла, характерное для Rhipineoptera: жилки Cu2 и Pcu (т.е. жилки, обрамляющие клавальный желоб и ограничивающие анальный веер спереди) на заднем крыле более длинные и прямые, чем на переднем (рис. 8.3.2D –E).
Возможны два объяснения того, что по строению анального веера Plecoptera отличаются от Tegminoptera: либо анальный веер, исходный для Rhipineoptera, у Plecoptera особым образом видоизменился, либо анальный веер разного строения независимо возник у Plecoptera и у Tegminoptera (см. Rhipineoptera: «Классификации» I).
(5) Утрата стерностилей. Стерностили [исходно на IX уромере – см. Pterygota (6)] полностью утрачены как у самца, так и у самки [о гениталиях самца см. (10)]. Неуникальная апоморфия (см. Указатель признаков [3.34] и [3.35]).
(6) Редукция яйцеклада. У самки первичный яйцеклад [см. Amyocerata (7)] утрачен, VIII и IX уромеры самки не имеют кинетапофизов (неуникальная апоморфия – см. Указатель признаков [3.51]). Несмотря на то, что яйцеклад утрачен у всех веснянок, строение вентральных стенок VIII и IX уромеров, образовавшихся на месте утраченных кинетапофизов, различается самок разных видов: VIII уромер либо имеет выраженный собственный стернит, либо не имеет стернита, так что на вентральной стороне брюшка VII стернит смыкается с IX стернитом; если стернит VIII уромера имеется, он может нести небольшие парные выросты, вероятно являющиеся вестигиями VIII кинетапофизов (рис. 8.3.1A). Генитальное отверстие самки первично находится на границе VII и VIII уромеров (Brinck 1956); у разных веснянок оно располагается либо сразу позади VII стернита, либо на VIII стерните (рис. 8.3.1A), либо на границе VIII и IX стернитов брюшка. Возможно, разнообразие в строении вентральной стороны генитальных сегментов и положения полового отверстия связано с тем, что редукция яйцеклада происходила независимо в разных таксонах веснянок.
Большинство веснянок откладывают яйца в полете, сбрасывая яйцевую массу в воду, либо смывают яйца в потоке.
(7) Строение гонад. Гонады часто имеют характерное строение: их передние слепые концы (исходно являющиеся парными) срослись между собой дорсальнее кишечника [тогда как их задние концы слиты в непарный проток вентральнее кишечника – см. Hexapoda (12)]; таким образом гонада образует замкнутую петлю. Такое строение имеют семенники самца и/или оварии самки многих видов, тогда как у части видов семенники и/или оварии имеют обычное строение, т.е. со свободными передними концами. У некоторых видов пара семенников, помимо того, что срастается своими передними концами, срастается также концами своих семенных пузырьков (каждый из которых исходно представляет собой слепой отросток, отходящий от задней части семенного протока), так что получается не одна, а две замкнутые петли, лежащие одна под другой (Brinck 1956). Срастание гонад в петлю характерно для многих веснянок и считается исходным для Plecoptera (Zwick 1973, 2000).
Плезиоморфии и изменчивые признаки Plecoptera
(8) Строение антенн. Флагеллум антенны у зрелой личинки и имаго примитивной щетинковидной формы с большим неопределенным числом члеников, без постоянного апикального членика; с каждым возрастом число члеников добавляется путем деления проксимальных члеников, при этом дистальные членики могут отбрасываться (рис. 8.3.2B–C) [см. Amyocerata (1c)]. У личинок первого возраста флагеллум обычно 7-члениковый (т.е. антенны 9-члениковые), но у некоторых Antarctoplecoptera число члеников иное (Sephton & Hynes 1982) (см. Указатель признаков [1.27]: табл. B).
(9) Строение брюшка. Брюшко личинки и имаго состоит из 10 уромеров [см. Hexapoda (8)] следующего строения. I уромер обычно имеет лишь обособленный тергит, тогда как его стернит слит с заднегрудью или утрачен. У каждого из II–IX уромеров тергит и стернит могут быть разделены мембранами (рис. 8.3.1A, 8.3.2A), либо слиты в цельное кольцо; обычно на нескольких передних уромерах тергит и стернит разделены, а на нескольких задних уромерах слиты; во многих таксонах число сегментов со слитыми тергитом и стернитом у личинки больше, чем у имаго. Последний, десятый уромер имеет разнообразное строение: у Euholognatha его тергит не замкнут вентрально, так что парапрокты причленяются к заднему краю IX стернита (рис. 8.3.2A) [первичное состояние – см. Amyocerata (8)]; у Metaplecoptera десятый тергит замкнут вентрально и образует цельное кольцо, позади которого находятся парапрокты (см. «Классификации» III) (то же у некоторых других насекомых – см. Указатель признаков [3.22]).
(10) Гениталии самца. Копулятивные аппараты самцов разнообразны. Во всех случаях самец при копуляции находится на спине самки; самец подгибает брюшко (слева или справа от самки), так что вершина брюшка самца, оставаясь направленной тергитами дорсально, оказывается под генитальным отверстием самки [см. (6)].
Генитальный аппарат самца (симметричный или асимметричный) устроен таким образом, что направляет сперму (в жидком виде или заключенную в сперматофор) дорсально-вперед. У многих веснянок функцию проведения спермы выполняют модифицированные части десятого сегмента брюшка – либо парапрокты, либо эпипрокт. В последнем случае эпипрокт (расположенный дорсальнее анального отверстия) образует склеротизованный копулятивный отросток, загнутый дорсально-вперед, огибающий анальное отверстие сзади и способный смыкаться с генитальным отверстием [расположенным вентральнее анального отверстия между IX и X уромерами – см. Amyocerata (6)]; на своей задней стороне этот копулятивный отросток имеет желоб, по которому сперма перекачивается из полового отверстия к дорсальному концу копулятивного отростка. У большинства Subulipalpia генитальная камера [находящаяся на вентральной стороне брюшка между IX и X уромерами – см. Amyocerata (6)] образует выпячивающийся пенис, который в выпяченном состоянии огибает вершину брюшка сзади и направлен дорсально-вперед. Церки либо не участвуют в образовании полового аппарата, либо преобразованы в хватательные придатки [см. (11)].
Обычно пара семенных протоков сливается в короткий непарный проток, открывающийся на вершине пениса или в генитальной камере непарным отверстием [как у большинства насекомых – см. Hexapoda (12)], но у некоторых Perlodes/fg семенные протоки на всем протяжении парные и открываются парой отверстий на вершине пениса (Brinck 1956).
(11) Разнообразие церков. Церки [только парные – см. Metapterygota (4)] у личинки и имаго часто длинные, многочлениковые; у личинок церки всегда многочлениковые, но у имаго многих Euholognatha церки короткие, одночлениковые, у самцов превращены в специализированные хватательные придатки копулятивного аппарата [см. (10)]. При линьке дистальные членики отбрасываются; это происходит и при тех линьках, при которых общее число члеников увеличивается путем деления проксимальных члеников [см. Amyocerata (8)].
Высказывалось утверждение, что у Plecoptera, помимо пары церков, бывает развит непарный парацерк, что рассматривалось как симплезиоморфия Plecoptera и Ephemeroptera (Illies 1960b). При этом за парацерк ошибочно принимали один (медиальный) из пяти или семи жаберных отростков, имеющихся на конце брюшка у личинок Austroperla/fg [см. (1Dd)] (рис. 8.3.3) (Illies 1969).
Размеры. Длина тела 3–40 мм.
Геологический возраст. Ранняя пермь – ныне (рис. 2.2). Наиболее древние веснянки известны из ранней перми Урала. Из ранней перми Урала описаны Uralonympha varica Zalessky 1939, Sylvoperlodes zhiltzovae Sinitshenkova 1987, Rasilopsis irrita Sinitshenkova 1987, Barathronympha victima Sinitshenkova 1987 и несколько видов, относимых к родам Perlopsis Martynov 1940 и Tshekardoperla Sinitshenkova 1987. Начиная с поздней перми веснянки известны и из Северного, и из Южного полушарий (Синиченкова 1987). Пермские представители Plecoptera известны как по взрослым насекомым, так и по личинкам; в их внешнем строении, доступном для изучения, не обнаружены признаки, которые бы не укладывались в характеристику Plecoptera, составленную по современным представителям.
Предположительно близким к Plecoptera считают Gulou carpenteri Béthoux et al. 2011 из карбона Китая; это насекомое похоже на веснянок, но отличается тем, что на заднем крыле жилка RS отходит не от MA, а от RA.
Состав и распространение. Около 4000 видов (Zhang 2011); во всех частях света, преимущественно в умеренных широтах, т.е. в Голарктике и Нотогее.
Систематическое положение Plecoptera. Кроме принятого здесь отнесения Plecoptera непосредственно к Rhipineoptera предлагались следующие классификации.
I. Объединение Plecoptera, Ephemeroptera и Odonata в таксон Arkiptera (см. Ephemeroptera: «Систематическое положение» III).
II. Объединение Plecoptera и Embioptera в таксон Plecopteroidea (см. Embioptera: «Систематическое положение» IV).
III. Объединение Plecoptera, Embioptera и Dermatoptera в таксон Panplecoptera (см. Embioptera: «Систематическое положение» V).
IV. Объединение Plecoptera, Embioptera, Isoptera и Copeognatha в таксон Platyptera (см. Embioptera: «Систематическое положение» III).
V. Объединение Plecoptera, Dermatoptera, Acercaria и Metabola в таксон Promeroista Büning 1996. Предполагаемой апоморфией этого таксона является неполное дихотомическое деление цистобластов в яичнике [см. Hexapoda (12)]. В яичнике Plecoptera все получающиеся при этом делении цистоциты в дальнейшем разделяются до конца и развиваются в яйцеклетки, так что трофоциты не образуются и яичники остаются паноистическими. У Dermatoptera, Acercaria и Metabola (объединяемых в таксон Meroista – см. Dermatoptera: «Систематическое положение» I) такое деление цистобласта обычно приводит к образованию кластера из трофоцитов и яйцеклетки. Однако среди Plecoptera деление цистобластов обнаружено лишь у одного исследованного вида Nemoura sp., тогда как большинство других насекомых с паноистическими яичниками в этом отношении вообще не исследованы (Büning 1996).
Классификации Plecoptera.
I. Деление Plecoptera на Holognatha и Systellognatha. Таксон Holognatha Enderlein 1909a {nom.typ.: Nemoura/fg [f:1853; g:1796] (incl. Eusthenia)} является плезиоморфоном и объединяет веснянок, у которых ротовой аппарат функционирует не только у личинок, но и имаго. Таксон Systellognatha Enderlein 1909a {nom.typ.: Perla/fg [f:1802; g:1762]} характеризуется нефункционирующим ротовым аппаратом у имаго (см. III) .
II. Деление Plecoptera на Arctoperlaria и Antarctoplecoptera. Таксон Arctoperlaria Zwick 1973 {nom.typ.: Perla/fg [f:1802; g:1762] (incl. Nemoura)} объединяет всех веснянок Арктогеи и родственные им таксоны Нотогеи. Центром многообразия Arctoperlaria является Голарктика (умеренная зона Северного Полушария), тогда как в тропиках и в умеренной зоне Южного Полушария представлены только Notonemoura/fg и отдельные таксоны, относящиеся к Perla/fg (sine Perlodes, Chloroperla). Единственной предполагаемой апоморфией Arctoperlaria является способность к звуковой коммуникации, при которой самец стучит брюшком по субстрату; однако морфологические приспособления для этого существенно различаются у разных таксонов, относимых к Arctoperlaria. Сходная звуковая коммуникация явно независимо возникла у разных насекомых (в частности, у ручейников – Trichoptera), так что этот признак недостаточен для обоснования голофилии Arctoperlaria.
Таксон Antarctoplecoptera (называвшийся «Antarctoperlaria» sensu Zwick 1973) объединяет веснянок нотогейского происхождения (см. III).
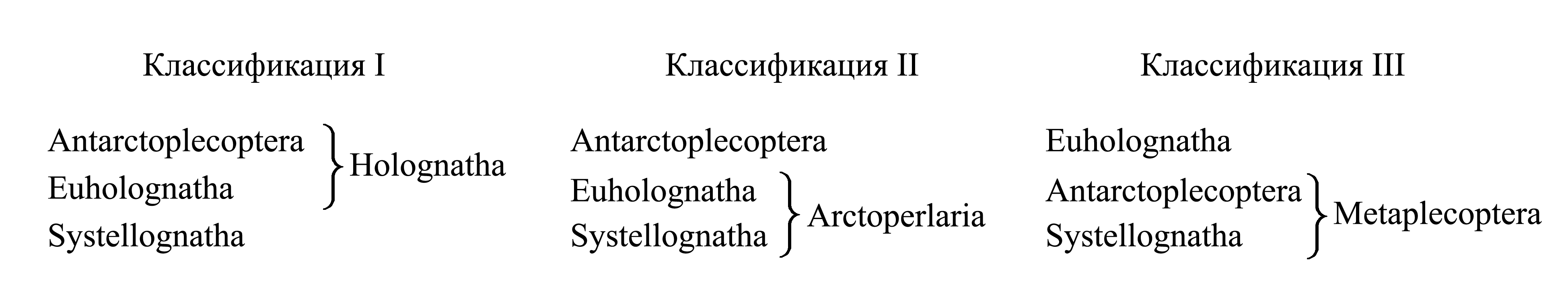
III. Деление Plecoptera на Euholognatha и Metaplecoptera. Здесь Antarctoplecoptera и Systellognatha объединены в новый таксон Metaplecoptera, который характеризуется вентральным слиянием десятого тергита брюшка личинки и противопоставляется таксону Euholognatha.
1. Плезиоморфон Euholognatha Zwick 1973 {nom.hier.: Nemoura/fg1 [f:1853; g:1796] (incl. Scopura)}. В отличие от Metaplecoptera, у личинки и имаго десятый тергит брюшка не сомкнут вентрально и не отделяет парапрокты от девятого уромера (рис. 8.3.1A, 8.3.2A) [см. Plecoptera (12)]. В отличие от Metaplecoptera-Systellognatha, у имаго сохраняется функционирующий ротовой аппарат, сходный с личиночным; личинки и имаго детритофаги с неспециализированным ротовым аппаратом.
1.1. Scopura/fg [fg:1935]. Личинка с жаберными нитями на X уромере и на границе IX и X уромеров; способна втягивать десятый уромер вместе с жабрами [см. Plecoptera (1Dc)]. 5 бескрылых видов в восточной Азии.
1.2. Filipalpia Klapalek 1905 (s.str.) {nom.hier.: Nemoura/fg2 [f:1853; g:1796] (incl. Taeniopteryx, Capnia, Leuctra)}. Включает около половины всех видов веснянок.
1.2.1. Taeniopteryx/fg1 [f:1905; g:1841] (incl. Brachypteryx). В отличие от прочих веснянок, у личинки и имаго второй членик лапки не укорочен, так что все три членика сходны по длине (рис. 8.3.1A, 8.3.2A). Около 100 видов в Голарктике.
1.2.2. Capnia/fg1 [f:1900; g:1841]. Характеризуется редукцией поперечных жилок на переднем крыле между M, Cu1 и Cu2 [см. Plecoptera (3)]. Более 200 видов в Голарктике.
1.2.3. Leuctra/fg1 [f:1905; g:1836]. Более 200 видов в Голарктике.
1.2.4. Nemoura/fg3 [f:1853; g:1796] (incl. Notonemoura). Крылья с характерной X-образной фигурой, состоящей из жилок между продольными жилками C, Sc, RA и RS у места разветвления RS. Около 750 видов в Голарктике, Ориентальной области и в Южном Полушарии (Южная Америка, Африка, Австралия, Тасмания, Новая Зеландия).
2. Metaplecoptera taxon n. {nom.hier.: Perla/fg2 [f:1802; g:1762] (sine Nemoura; incl. Eusthenia); по циркумскрипции соответствует Perlina: Синиченкова 1987}. У личинки и самки имаго десятый тергит брюшка сомкнут вентрально, полностью отделяя находящиеся позади него парапрокты от девятого сегмента брюшка (рис. 8.3.3) [см. Plecoptera (12)]. Авторы, считающие брюшко насекомых исходно 11-сегментным, рассматривают этот признак как плезиоморфию, поскольку принимают десятый тергит брюшка за весь десятый уромер, а парапрокты принимают за компоненты одиннадцатого уромера (Zwick 2000). В действительности парапрокты принадлежат десятому уромеру; об этом свидетельствует то, что у веснянок, как и у других насекомых, пара вентральных мышц, идущих к основаниям парапроктов, начинается от переднего края девятого стернита брюшка (рис. 8.3.3: v9-10), а мышцы, идущие к основаниям церков, начинаются от десятого тергита брюшка [см. Amyocerata (8)]. В отличие от личинок и самок, у взрослых самцов Metaplecoptera десятый тергит брюшка может быть не сомкнут вентрально, что связано с его преобразованием в копулятивный аппарат [см. Plecoptera (10)].
2.1. Antarctoplecoptera taxon nov. {nom.hier.: Gripopteryx/f1=Eusthenia/g1 [f:1909; g:1832]; по циркумскрипции соответствует «Antarctoperlaria» sensu Zwick 1973}. Название «Antarctoperlaria» является типифицированным названием с основой Antarctoperla/f,; применительно к данному таксону это название было употреблено без соблюдения принципа приоритета: исходно семейство Gripopterygidae делилось на подсемейства Gripopteryginae и Antarctoperlinae (Enderlein 1909), так что Gripopteryx/f [f:1909] имеет приоритет над Antarctoperla/f [f:1909]. Предположительной аутапоморфией является модификация в мускулатуре передних ног (Zwick 1973). В отличие от Systellognatha, у имаго сохраняется функционирующий ротовой аппарат, сходный с личиночным; личинки и имаго детритофаги или хищники с неспециализированным ротовым аппаратом. Имеют почти исключительно амфинотическое распространение – Южная Америка (преимущественно Чили), Австралия, Тасмания и Новая Зеландия; лишь один неописанный вид, относящийся к Gripopteryx/f2=Letoperla/g1, обнаружен на юге Индии.
2.1.1. Gripopteryx/f2=Leptoperla/g1 [f:1909; g:1839]. У личинки на вершине десятого сегмента брюшка дорсальнее анального отверстия имеется втягивающийся пучок жаберных нитей [см. Plecoptera (1Dd)]. Более 100 видов, распространенных в Южной Америке, Австралии, Тасмании и Новой Зеландии. Один неописанный вид в Индии.
2.1.2. Austroperla/fg1 [f:1921; g:1905]. У личинки имеются невтягивающиеся жаберные отростки на церках, парапрокте и эпипрокте (рис. 8.3.3) [см. Plecoptera (1Dd)]. 15 видов, распространенных на юге Южной Америки (Чили и юг Аргентины), в Австралии, Тасмании и Новой Зеландии.
2.1.3. Eusthenia/f1=g2 [f:1921; g:1832]. У личинки имеются нитевидные членистые парные жаберные отростки на первых 5 или 6 сегментах брюшка [см. Plecoptera (1Dc]. Более 20 видов на юге Южной Америки, в Асвстралии и в Новой Зеландии.
2.1.4. Diamphipnoa/fg1 [f:1950; g: 1873]. У личинки имеются подвижные кустистые парные жабры на I–IV сегментах брюшка [см. Plecoptera (1Dc]. Несколько видов в Чили и соседних районах Аргентины.
Иногда таксоны Eusthenia/fg и Diamphipnoa/fg объединяют в таксон Archiperlaria Illies 1960a.
2.2. Systellognatha Enderlein 1909a {nom.hier.: Perla/fg3 [f:1802; g:1762] (incl. Pteronarcys, Peltoperla)}. У имаго ротовой аппарат нефункционирующий, мандибулы обычно тонкие и слабые. Исходно в подотряд Systellognatha был включен род Diamphipnoa, ошибочно отнесенный к семейству Pteronarcidae (Enderlein 1909a), а в настоящее время относимый к Antarctoplecoptera (см. выше).
2.2.1. Pteronarcys/fg [f:1853; g:1838]. Более 10 видов в Восточной Палеарктике и Неарктике.
2.2.2. Peltoperla/fg [f:1931; g:1905]. Около 100 видов в Азии и Северной Америке.
2.2.3. Subulipalpia Klapalek 1905 {syn.circ.: Setipalpia Klapalek 1909; nom.hier.: Perla/fg4 [f:1802; g:1762] (incl. Perlodes, Chloroperla)}. Ротовой аппарат личинок специализирован для хищничества: мандибулы без молы и с удлиненным инцизором; глоссы нижней губы значительно меньше параглосс. При этом у имаго ротовой аппарат обычно нефункционирующий [см. Systellognatha].
2.2.3.1. Perlodes/fg [f:1909; g:1903]. Плезиоморфон в составе Subulipalpia. Более 200 видов в Голарктике.
2.2.3.2. Chloroperla/fg [f:1912; g:1836]. Анальный веер заднего крыла редуцирован [см. Plecoptera (4)]. Более 100 видов в Голарктике.
2.2.3.3. Perla/fg5 [f:1802; g:1762] (sine Perlodes, Chloroperla). Личинки с кустистыми жабрами на вентральной стороне груди [см. Plecoptera (1D)]. Включает около половины всех видов веснянок; широко распространены в Голарктической, Неотропической, Ориентальной и Афротропической областях.
* * *