КЛАДОЭНДЕЗИС НАСЕКОМЫХ

![]()
|
КЛАДОЭНДЕЗИС НАСЕКОМЫХ |
|||||||
|
|
|
fg | |||||
VIII-1.2.2.4. Dermatoptera Burmeister 1838 – уховертки
(Somobialia![]() Eucaryota
Eucaryota![]() Metazoa
Metazoa![]() Monostomata
Monostomata![]() Haemataria
Haemataria![]() Peritoneata
Peritoneata![]() Metameria
Metameria![]()
Gnathopoda![]() Euarthropoda
Euarthropoda![]() Mandibulata
Mandibulata![]() Atelocerata
Atelocerata![]() Hexapoda
Hexapoda![]() Amyocerata
Amyocerata![]()
Pterygota![]() Metapterygota
Metapterygota![]() Neoptera
Neoptera![]() Rhipineoptera
Rhipineoptera![]() Tegminoptera
Tegminoptera![]() Dermatoptera)
Dermatoptera)
(рис. 8.7.1–8.7.3)
(рис. 8.7.1,
8.7.2, 8.7.3)
|
Названия, соответствующие по циркумскрипции Dermapteroidea, Brachydermaptera, Neodermaptera и Holodermaptera: — Labidoura Dumeril 1805 (исходно только во французском написании Labidoures и в немецком написании Labidouren); — Placoda Billberg 1820; — Trimera Zetterstedt 1821 (non Trimera Dumeril 1805); — «Euplekoptera» Westwood 1831 (с опечаткой); — Euplexoptera Westwood 1838; — Dermatoptera Burmeister 1838; — Labidura (Burmeister 1838) Haeckel 1866 (non genus Labidura Leach 1815); — Dermoptera Agassiz 1848; — Harmoptera Fieber 1852; — Euplectoptera Fischer 1853 (non Euplectoptera Tillyard 1932); — Pseudoorthoptera Dominique 1893 (исходно только во французском написании Pseudoorthoptères). Названия, соответствующие по циркумскрипции Dermapteroidea, Brachydermaptera и Neodermaptera: — Dermaptoria Blatchley 1920. По циркумскрипции также соответствует: — «Dermaptera» sensu Kirby 1813 (non De Geer 1773); — таксонам под родовым названием Forficula (sensu Linnaeus 1758) и производными типифицированными названиями: Forficulariae, Forficulaedes, Forficulidae, Forficularia, Forficulites, Forficulina, Forficulodea, Forficulida, Forficulidi. Иерархическое название: Forficula/(f1–4)=(g3–6) [f:1810; g:L.1758(gen.192)]. |
|
Ранг. Отряд или надотряд.
Названия. Для этого таксона часто употребляют название «Dermaptera» с авторством «De Geer 1773». В действительности исходно таксон Dermaptera De Geer 1773 {syn.circ.: Ulonata Fabricius 1775} включал роды Mantis, Locusta, Acrydium, Gryllus, Blatta и Forficula (что соответствует нынешним таксонам Raptoriae, Spectra, Saltatoria, Neoblattariae и Dermatoptera) (см. Tegminoptera: «Название»); Позже Ретзиус (Retzius 1783), использовавший классификацию Де Геера, переименовал Dermaptera в Hemiptera (о разных значениях названия Hemiptera см. ниже, Arthroidignatha: «Название»), а название Dermaptera употребил для таксона, ныне называемого Heteroptera (см. ниже). Kirby (1813) предложил употреблять название Dermaptera применительно только к уховерткам (что соответствует только роду Forficula в классификации Де Геера). Для того, чтобы соблюсти циркумскриптный принцип номенклатуры, Westwood (1831) предложил для уховерток новое название Euplexoptera. Другим названием, соответствующим по циркумскрипции, является Dermatoptera Burmeister 1838; несмотря на то, что это название младше, оно мне кажется более удобным, т.к. сходно с широко известным, но неверным названием «Dermaptera»; название Euplexoptera сходно с названием Euplectoptera Tillyard 1932, используемым в настоящее время для поденок.
Наиболее старое название Labidoura (или Labidura) Dumeril 1805 нежелательно использовать в качестве циркумскриптного, поскольку оно является гемиомонимом родового названия Labidura Leach 1815. Название Placoda Billberg 1820 было забыто и не используется.
Если рассматривать только тех представителей Dermatoptera, которые были известны науке до 70-х годов XIX века, то таксон Dermatoptera выглядит четко очерченным. В этом случае все названия, перечисленные выше, однозначно соответствуют по циркумскрипции. Однако позже были описаны аберрантные вторичнобескрылые Diploglossata, а также вымершие палеозойские и мезозойские крылатые насекомые, которые имеют явное родство с настоящими уховертками, но не укладываются в таксон Dermatoptera, охарактеризованный только по строению известных ранее представителей. Эти формы вместе с настоящими уховертками образуют голофилетическую группу Dermatoptera s.1., или Dermapteroidea (или Forficula/f1=g2), внутри которой находится подчиненный таксон Dermatoptera s.2, или Brachydermaptera (или Forficula/f2=g3); внутри Brachydermaptera находится подчиненный таксон, объединяющий всех современных уховерток – Dermatoptera s.3, или Neodermaptera (или Forficula/f3=g4). Мы не имеем возможности рассмотреть все интересующие нас признаки этих соподчиненных таксонов, так как для вымерших форм известны лишь немногие детали строения, а для Diploglossata неизвестно исходное строение крыльев. Поэтому здесь характеристика уховерток разбита на две части. В первой части – «А. Общая характеристика Dermatoptera» рассматриваются признаки уховерток, про многие из которых неясно, относятся ли они ко всем Dermatoptera в широком смысле, или лишь к Dermatoptera в одном их более узких значений слова. Во второй части – «B. Классификация Dermatoptera с учетом вымерших групп» рассматриваются признаки, распределение которых в этих группах достоверно известно.
VIII-1.2.2.4.A. Общая характеристика Dermatoptera
Аутапоморфии Dermatoptera. О некоторых их этих признаков неясно, являются ли они аутапоморфиями Dermapteroidea, Brachydermaptera или Neodermaptera, так как для ископаемых групп Archidermaptera и Protelytroptera они неизвестны.
(1) Утрата глазков. Простые глазки отсутствуют [неуникальная апоморфия – см. Amyocerata (4)]. Иногда за глазки ошибочно принимают места прирастания дорсальных рук тенториума ко лбу между глазами. Глазки были указаны для вымершего таксона Archidermaptera и изображены на рисунках типовых экземпляров Archidermapteron martynovi и Semenovioloides capitatus (Вишнякова 1980); в действительности у этих экземпляров глазков не видно, и неизвестно, были ли глазки у Archidermaptera. Фасеточные глаза обычно нормально развиты, небольшие; реже отсутствуют.
(2) Подотельсон. Дистальный членик каждого щупика [т.е. 5-й членик максиллярного щупика и 3-й членик лабиального щупика – см. [см. Hexapoda (2c), (3c) и Rhipineoptera (3)] несет на вершине подотельсон (podotelson) – маленький цилиндрический выступ с сенсиллами на вершине; таким образом, в строении щупиков проявляется вторичная эволюционная корреляция (см. I.2.3). У 5-членикового максиллярного щупика два проксимальных членика укорочены, так что он кажется трехчлениковым; у 3-членикового лабиального щупика первый членик укорочен, так что он кажется двучлениковым. Это свойственно по крайней мере Neodermaptera; для вымерших групп строение щупиков неизвестно.
(3) Утрата глосс. Глоссы нижней губы [см. Hexapoda (3)] утрачены, так что имеется лишь одна пара апикальных придатков – хорошо развитые параглоссы. Это свойственно по крайней мере Neodermaptera; для вымерших групп строение нижней губы неизвестно. Встречающееся в литературе указание на наличие у уховерток непарной глоссы является ошибочным: в этом случае за глоссу принимают гипофаринкс.
(4) Вентральные шейные склериты. Помимо пары латеральных шейных склеритов [см. Pterygota (2)], имеются два подвижно соединенных вентральных шейных склерита – передний и задний (рис. 8.7.1A). У всех уховерток голова занимает прогнатное положение; при этом, в отличие от некоторых других насекомых с прогнатной головой, у уховерток головная капсула не распространяется на вентральную сторону, гула не образуется, а задний тенториальный мост, максиллы и нижняя губа сохраняют исходное положение и не смещены вперед. При таком строении головы вентральная сторона шеи удлинена, а вентральные шейные склериты защищают ее от повреждений. Различают два типа шейных склеритов: пигидикраноидный, при котором оба вентральных шейных склерита имеют вид небольших поперечных полос, и форфикулоидный, при котором задний вентральный шейный склерит увеличен и занимает значительную часть вентральной поверхности шеи; пигидикраноидный тип присущ надсемейству Pygidicranoidea, которое принято считать наиболее примитивным среди Neodermaptera, а форфикулоидный тип присущ большинству уховерток (см. ниже, Neodermaptera: «Классификации» IV).
(5) Модификации крылового аппарата. Крыловой аппарат [см. Tegminoptera (1)–(3)] сильно видоизменен: задние крылья имеют уникальную модификацию анального веера, позволяющую им складываться не только веером, но и поперек [см. (5a)], передние крылья имеют уникальный аппарат, позволяющий фиксировать их на спине [см. (5b)]; птероторакс имеет характерную форму [см. (5c)]. Протоптероны личинки имеют особенности, связанные с формой птероторакса и имагинальных крыльев [см. (5d)]. Помимо крылатых видов, среди уховерток есть короткокрылые и бескрылые; особенности их строения связаны со строением имагинальных крыльев и личиночных протоптеронов у исходных крылатых форм [см. (5e)].
(5 a) Задние крылья. На заднем крыле инверталярная область [образующая анальный веер – см. Tegminoptera (3)] модифицирована уникальным образом: она длиннее, чем лежащая впереди нее непереворачивающаяся область, так что вершинная часть крыла несет только веерообразно расположенные ваннальные жилки; эта вершинная часть крыла с ваннальными жилками способна складываться, сгибаясь вперед; проксимальные части ваннальных жилок слиты друг с другом таким образом, что анальный веер расходится не от основания крыла, а имеет вид либо гребенки (у Protelytroptera) (рис. 8.7.2F), либо веера, расходящегося из точки, сильно удаленной от основания крыла (у Brachydermaptera) (рис. 8.7.1A) [см. ниже, Dermapteroidea (2), Protelytroptera (1) и Brachydermaptera (1)].
У всех современных крылатых уховерток заднее крыло имеет следующее строение (рис. 8.7.1). Область крыла впереди анального веера преобразована в так называемый фустис (fustis): он сильно уменьшен, склеротизован, с обедненным жилкованием. У полнокрылых форм при сложенных крыльях дистальная часть фустиса всегда выступает сзади из-под короткого надкрылья [см. (5b)] (рис. 8.7.2C). Ваннальные жилки идут от основания крыла общим изогнутым стволом, который огибает сзади фустис и дистально причленяется к вершине фустиса; от места сочленения общего ваннального ствола с вершиной фустиса веером расходятся 8 ваннальных жилок (V1 –V8), каждая из которых своим основанием подвижно причленена к вершине общего ствола ваннальных жилок; передние из этих восьми ваннальных жилок в расправленном виде направлены дистально, так что крыло с расправленными ваннальными жилками значительно длиннее фустиса. Позади группы из восьми ваннальных жилок от общего ваннального ствола отходят порознь еще две ваннальные жилки (V9 –V10). Между каждыми двумя соседними ваннальными жилками находится интеркалярная жилка, имеющая свободное основание. Таким образом, весь анальный веер состоит из 10 ваннальных и 9 интеркалярных жилок. Все ваннальные и интеркалярные жилки соединены одной цепью поперечных жилок; каждая из этих поперечных жилок имеет слабый участок, по которому при складывании веера проходит радиальная складка. Проксимальнее цепи поперечных жилок каждая ваннальная и интеркалярная жилка имеет уплощенный и расширенный участок характерной изогнутой формы; у ваннальных и интеркалярных жилок изгибы уплощенных участков направлены в противоположные стороны. При складывании крыла анальный веер складывается так, что все интеркалярные жилки переворачиваются; сложенный веер сгибается пополам по линии, проходящей по уплощенным расширенным участкам жилок; это происходит в результате того, что по мере отведения жилок назад их дистальные участки отгибаются вперед. Таким образом сложенное крыло оказывается по длине равно его фустису и может быть в три раза короче расправленного крыла и в два раза короче каждой ваннальной жилки.
Способ складывания заднего крыла тот же, что при его складывании в процессе роста под кутикулой личиночного протоптерона [см. (5d)] (рис. 8.7.2A), так что сложенное положение является устойчивым. Для расправления крыльев перед полетом уховертка использует брюшко и церки.
У некоторых нелетающих уховерток задние крылья сильно уменьшены, но при этом сохраняется их строение, число жилок и способность складываться.
У вымерших Archidermaptera жилкование крыльев неизвестно, т.к. найдены только экземпляры со сложенными крыльями. Поскольку у них дистальные части фустисов выступают из-под надкрыльев так же, как у современных уховерток (рис. 8.7.2C), можно предположить, что и все строение задних крыльев у них было таким же, как у современных уховерток – т.е. описанные выше особенности строения крыльев являются общими для Brachydermaptera. В отличие от них, у Protelytroptera строение заднего крыла несколько более примитивное [см. ниже, Protelytroptera (1) и рис. 8.7.2F].
(5 b) Передние крылья и аппарат их фиксации. Передние крылья [имеющие тенденцию к элитризации – см. Tegminoptera (2)] превращены в плотные надкрылья (тегмины). В отличие от сходных по внешнему виду элитр жуков, тегмины уховерток образованы не жилками, а утолщенной склеротизованной крыловой мембраной, так что состоят из двух слоев кутикулы, сомкнутых по всей площади тегмена. Широкая полоса вдоль костального края надкрылья образует эпиплевру – область надкрылья, расположенную под прямым углом к остальной поверхности надкрылья так, что и при сложенных надкрыльях она налегает сбоку на плейрит [см. Neoptera (1c)]; благодаря сильному наклону птероторакса [см. (5с)] эпиплевра закрывает дорсальную часть плейрита, начиная от переднего края сегмента. Анальный край надкрылья всегда почти прямой; в покое надкрылья всегда сложены на спине так, что их анальные края сходятся и налегают друг на друга лишь узкими полосками. Такое постоянное положение сложенных надкрыльев обеспечивается особым фиксирующим приспособлением (рис. 8.7.1C): на вентральной стороне каждого надкрылья вдоль его анального края (который при сложенных крыльях является медиальным) тянется продольный гребень, несущий частые крепкие щетинки, направленные латерально-назад; на скутуме заднегруди имеется непарное продольное медиальное вдавление, обрамленное парой продольных гребней, каждый из которых несет частые крепкие щетинки, направленные медиально-назад. Такое приспособление для фиксации передних крыльев имеется не только у крылатых видов, но и у большинства видов, утративших задние крылья, но сохранивших надкрылья [см. (5e)]. У личинок метанотум не имеет ни медиального вдавления, ни щетинистых гребней; то же у бескрылых имаго с ларвализованным нотумом [см. (5e)]. Судя по некоторым ископаемым остаткам, это фиксирующее приспособление имеется не только у современных уховерток, но и у ископаемых Protelytroptera и Archidermaptera, т.е. исходно присуще Dermapteroidea.
У вымерших Protelytroptera надкрылья закрывают все брюшко, у Brachydermaptera (куда входят все современные уховертки) надкрылья сильно укорочены; жилкование надкрыльев сохраняется у вымерших Protelytroptera и Archidermaptera (рис. 8.7.2B, C, E) и утрачено у всех современных уховерток [см. ниже, Dermapteroidea (1), Brachydermaptera (1) и Neodermaptera (3)].
(5 c) Плейриты груди. Плейриты всех трех сегментов груди сильно наклонены дорсальной стороной вперед [см. Neoptera (2)]; на переднегруди дорсальный конец плейрального шва находится у самого переднего края пронотума, так что эпистернум не имеет границы с пронотумом; основания передних крыльев находятся на самом переднем крае мезонотума; основания задних крыльев находятся на самом переднем крае метанотума (рис. 8.7.1A). Благодаря этому сложенные надкрылья закрывают весь мезонотум, а у личинки передние протоптероны совпадают с мезонотумом [см. (5d)] (рис. 8.7.2A). В связи с наклонной формой груди первый стернит брюшка утрачен (неуникальный признак – см. Указатель признаков [3.9]).
Сходная форма плейритов средне- и заднегруди свойственна Idioprothoraca, но форма плейритов переднегруди у Idioprothoraca иная.
(5 d) Протоптероны личинки. У личинок крылатых уховерток протоптероны выражены только в последних возрастах и имеют характерное строение, из-за которого передние протоптероны кажутся отсутствующими (рис. 8.7.2A). И передние, и задние протоптероны паннотные. Передние протоптероны, будучи пропорциональны крыльям, у современных уховерток короткие [см. (5b)]; из-за сильного наклона плейритов [см. (5c)] они начинаются от самого переднего края среднегруди, так что их вершины не выступают за задний край мезонотума; эпиплевры протоптеронов, как и эпиплевры сложенных имагинальных крыльев, налегают на плейриты, закрывая дорсальную часть плейрита начиная от самого переднего края сегмента. Мезонотум вместе с передними протоптеронами выглядит как цельный тергит с лопастевидными латеральными краями, загнутыми вниз и закрывающими дорсальные части плейритов. При развитии передних крыльев под кутикулой личинки, они заполняют латеральные части и заднюю часть тергита, смыкаясь медиально (рис. 8.7.2A).
Протоптероны задних крыльев, в отличие от передних, в последнем возрасте отчетливо выступают назад от метанотума, так что кажутся единственной парой протоптеронов. Они имеют форму и рисунок, соответствующие уникальной форме и жилкованию расправленного заднего крыла [см. (5a)]. При развитии заднего крыла под кутикулой протоптерона оно складывается по тем же складкам, что и сложенное крыло имаго (рис. 8.7.2A), и сминается по многочисленным дополнительным складкам; при выходе имаго из личиночного экзувия дополнительные складки расправляются, и имагинальное крыло сразу оказывается в сложенном положении.
(5 e) Бескрылые формы. Как и среди многих других насекомых, среди Dermatoptera имеются бескрылые виды, имаго которых имеют либо вестигии имагинальных крыльев, либо вестигии личиночных протоптеронов [см. Neoptera (1e)].
В первом случае обычно сохраняются вполне развитые надкрылья с фиксирующим приспособлением [см. (5b)], а задние крылья либо сильно уменьшены, но сохраняют присущее им жилкование и способность складываться [см. (5a)], либо утрачены.
Во втором случае мезонотум с вестигиями передних крыльев выглядит так же, как мезонотум личинки с протоптеронами [см. (5d)] – т.е. как цельный тергит с лопастевидными латеральными краями, загнутыми вниз и закрывающими дорсальные части плейритов. В отличие от личинок крылатых видов, у таких бескрылых уховерток вестигии задних крыльев такие же короткие, как вестигии передних.
(6) Утрата шпор. Шпоры на вершине голени [см. Neoptera (4)] отсутствуют. У некоторых уховерток на месте шпор имеются короткие крепкие щетинки. У Allostethus lombokianum Verhoeff 1904 на месте шпор имеются два наклонных шипа, каждый с сенсиллой у основания; в отличие от настоящей шпоры, такой шпоровидный шип при каждой линьке возникает заново, так что перед линькой лежит вне старого шипа; вероятно такой шип является результатом видоизменения кутикулярного возвышения, окружающего сочленовную мембрану сенсиллы [см. Euarthropoda (5)].
(7) Модификация прегенитальных сегментов брюшка. Задние края тергитов и стернитов брюшка широко налегают на позади лежащие тергиты и стерниты, а боковые края тергитов (параноты) налегают на стерниты; исключение составляют VIII–IX уромеры самки [см. (8)] (рис. 8.7.1A). В результате этого каждая пара абдоминальных дыхалец [имеющихся на плеврах I–VIII уромеров – см. Amyocerata (2)] закрыта паранотами своего сегмента, а поверх них – еще налегающими задними краями впереди лежащего стернита, так что снаружи дыхальца не видны. Подобное строение брюшка независимо возникло у щетинохвосток [ср.: Triplura (1)].
У самца наружная стенка брюшка состоит из 10 тергитов (I–X) и 8 видимых стернитов (II–IX), поскольку первый уростернит утрачен [см. (5c)], а вентральная сторона десятого уромера прикрыта девятым уростернитом [см. (9)]. У самки наружная стенка брюшка состоит из 8 видимых тергитов (I–VII и X) и 6 видимых стернитов (II –VII), поскольку VIII и IX уромеры скрыты [см. (8)] (рис. 8.7.1A, 8.7.3E). Вершина брюшка внешне кажется сходной у самца и самки, поскольку десятый сегмент увеличен [см. (10)] и с вентральной стороны прикрыт удлиненным стернитом предыдущего видимого сегмента, который является девятым у самца и седьмым у самки.
(8) Модификация генитальных сегментов самки. У имаго самки последние сегменты брюшка видоизменены [см. (8a)]; яйцеклад утратил олистетеры, а его первые створки могут быть усложнены и преобразованы для самостоятельного проведения яиц [см. (8b)].
(8 a) Сегменты брюшка самки. У самки VIII и IX уромеры [исходно несущие яйцеклад – см. Amyocerata (7)] сильно укорочены и неподвижно соединены с десятым уромером; тергиты VIII и IX уромеров имеют форму узких полуколец в основании цельного VIII–X уромера, разделенных более или менее выраженными бороздами; в результате этого дыхальца VIII уромера [последней пары – см. Amyocerata (2)] находятся на цельном VIII–X уромере (рис. 8.7.3F). Тергит и стернит VII уромера [см. (7)] полностью закрывают VIII и IX уромеры; при этом стернит VII уромера увеличен, простирается назад и с вентральной стороны налегает на X уромер (рис. 8.7.1A, 8.7.3E). Так что внешне брюшко самки кажется состоящим из 8 тергитов (II–VII и X) и 6 стернитов (II–VII) [поскольку I стернит редуцирован – см. (5c)]. По форме видимых сегментов брюшко самки сходно с брюшком самца, но ее видимый предпоследний уромер, стернит которого налегает на X уромер, является седьмым, тогда как у самца сходный по форме уромер является девятым (рис. 8.7.3B).
В отличие от имаго, у молодых личинок самок VIII и IX уромеры вполне развиты, подвижны и не скрыты, так что все уромеры равны по длине, как и у самцов.
Такое строение брюшка свойственно вымершим Archidermaptera (рис. 8.7.2B) и подавляющему большинству современных Neodermaptera. Однако у Plecodermaptera (см. ниже, Neodermaptera: «Классификации» I) восьмой и девятый уромеры самки не уменьшены, их тергиты не слиты с десятым тергитом и видны снаружи; вероятно, в ходе эволюции этих бескрылых насекомых произошла ювенилизация, в результате чего брюшко половозрелых особей стало похожим на брюшко личинок. В литературе имеется ошибочное указание, что у Archidermaptera VIII и IX уромеры самки не укорочены (Вишнякова 1980); это связано с ошибкой в подсчете видимых сегментов (установлено нами при переисследовании тех же экземпляров).
Помимо Dermatoptera, редукция VIII и IX уромеров имеет место и у некоторых других насекомых, в частности у некоторых короткокрылых и бескрылых Neoblattariae.
(8 b) Яйцеклад. В результате того, что у взрослых самок VIII и IX уромеры укорочены и скрыты, первичный яйцеклад [см. Amyocerata (7)] не может отгибаться вентрально и сохранять свои исходные функции. У вымерших мезозойских Archidermaptera яйцеклад выглядит как вполне развитый: длинный, прямой, состоит из сомкнутых створок (рис. 8.7.2B); однако детали его строения неизвестны [см. ниже, Archidermaptera (3)].
У всех современных уховерток яйцеклад утратил олистетеры; у большинства уховерток яйцеклад совсем утрачен (рис. 8.7.1A). У некоторых уховерток на вентральной стороне слитного VIII–IX уромера имеются небольшие плоские лопастевидные выросты, отходящие назад от мест, соответствующих задним краям VIII и/или IX уромеров; у разных видов эти лопасти различаются по форме, бывают парными и непарными; некоторые из парных лопастей представляют собою нефункционирующие вестигии VIII кинетапофизов, IX кинетапофизов и IX стилигеров (стерностили IX уромера, вероятно, полностью утрачены, и их вестигии не обнаружены). Все эти лопасти полностью или частично скрыты налегающим стернитом VII уромера.
У некоторых видов Pygidicrana/fg (см. Neodermaptera: «Классификации»), имеется вторичный функционирующий яйцеклад следующего строения (рис. 8.7.3E–F). VIII кинетапофизы направлены назад, внутри них проходят продольные мышцы (являющиеся новообразованиями). Каждый из этих кинетапофизов имеет утолщенную проксимальную часть и тонкую дистальную часть. У проксимальных частей медиальные стороны вогнутые, так что, смыкаясь вместе, они образуют канал для проведения яиц. Дистальные части кинетапофизов тоньше яйца; у некоторых видов они способны к самостоятельным движениям, т.к. вмещают продольные мышцы, заходящие в них из проксимальных частей, укреплены упругими склеротизованным продольными ребрами и настолько длинные, что выступают назад из-под VII стернита брюшка; у некоторых видов дистальные доли кинетапофизов вестигиальные – короткие, мягкие и лишены мышц. IX кинетапофизы иногда присутствуют в виде мягких нефункционирующих вестигиев, не сцеплены с VIII кинетапофизами. Таким образом, в яйцекладе определенной части уховерток вместо двух пар сцепленных кинетапофизов функцию проведения яиц выполняет одна пара кинетапофизов, которые приобрели усложненное строение и внутренние мышцы. Вероятно, такой яйцеклад, утративший способность поворачиваться вентрально, приспособлен для функционирования под плотно налегающим VII стернитом брюшка.
Модификация и редукция первичного яйцеклада у уховерток связана с заботой о потомстве: у изученных в этом отношении современных видов самка откладывает яйца в укрытии, где продолжает находиться до вылупления личинок.
(9) Копулятивный аппарат самца. Гениталии самца [расположенные на границе XI и X уромеров – см. Amyocerata (6)] имеют следующее строение (рис. 8.7.3B–D). Стернит IX уромера простирается далеко назад, так что закрывает снизу и копулятивный аппарат, и вентральную сторону десятого уромера [см. (7)]; стерностили IX уромера [исходно безмышечные – см. Metapterygota (5)] полностью утрачены. Копулятивный аппарат представляет собой удлиненную капсулу, которая соединена только с эластичной сочленовной мембраной между IX и X уромерами, так что способна втягиваться вперед, выдвигаться назад и поворачиваться; в покое она глубоко втянута вперед, заходя своим основанием в VIII уромер (рис. 8.7.3B). В своей дистальной (задней) части копулятивный аппарат несет пару латеральных придатков, называемых парамерами, или наружными парамерами, хотя они не гомологичны истинным парамерам жуков [см. Amyocerata (6) и Metabola (6)]. Парамеры направлены вершинами апикально (т.е. назад); они могут быть неподвижными и иметь простую удлиненную форму, а могут быть хватательными – склеротизованными, с зубцами на вершине и/или на медиальной стороне; к их основаниям могут подходить мышцы. Между парамерами находится один или два выроста – пенисы. От вершины пениса вглубь него вдается мембранозный мешок, соединенный с семяизвергательным каналом и способный выворачиваться и выдвигаться под действием находящихся в пенисе мышц; семяизвергательный канал на большом протяжении эктодермальный и выстлан кутикулой, а в дистальной части его кутикула образует виргу – склеротизованную трубку видоспецифичной формы.
Среди разнообразия копулятивных аппаратов уховерток можно выделить два основных типа – диандрический и монандрический.
Диандрический копулятивный аппарат (рис. 8.7.3C) имеет непарное основание и раздваивается дистально – т.е. имеет Y-образную форму; его дистальная часть образована парой пенисов, к каждому из которых латерально крепится один из пары парамеров. В типичном случае каждый пенис разделен на проксимальную часть, неподвижную по отношению к основанию копулятивного аппарата, и подвижную дистальную часть – телопенис (telopenis). В покое телопенисы лежат на дорсальной стороне копулятивного аппарата и направлены вершинами в сторону его основания (т.е. вперед по отношению к телу насекомого); в активном состоянии телопенисы поворачиваются на 180°, так что оказываются расположенными между парамерами и направлены вершинами дистально (т.е. назад по отношению к телу насекомого). В проксимальной части копулятивного аппарата проходит непарный семенной проток [как и у всех уховерток, он выходит из непарного резервуара, в который впадает пара семенных протоков, идущих от пары семенников – см. Hexapoda (12)]; дистальнее семенной проток раздваивается, так что в каждом из двух пенисов проходит свой проток; каждый из этих двух протоков в своей дистальной части образует склеротизованную виргу, заканчивающуюся половым отверстием; таким образом, имеется 2 половых отверстия. У некоторых видов Diplatys/fg и Pygidicrana/fg (см. Neodermaptera: «Классификации») каждая вирга, в свою очередь, дистально раздвоена, и в этом случае имеется в общей сложности 4 половых отверстия. В некоторых таксонах и у отдельных особей один из двух пенисов может быть нефункционирующим, а иногда совсем утрачен; в этих случаях имеется только одно половое отверстие.
Монандрический копулятивный аппарат (рис. 8.7.3D) имеет один пенис, постоянно направленный вершиной дистально (т.е. назад по отношению к телу насекомого) и расположенный между парамерами. Половой проток на всем протяжении непарный, несет одну виргу и открывается на вершине пениса одним половым отверстием.
Принято считать, что диандрический аппарат является исходным, а монандрический – производным от него (см. Neodermaptera: «Классификации» III). Это утверждение отчасти основано на мнении, что парный половой аппарат в принципе примитивнее непарного; в действительности же пенис насекомых исходно непарный [см. Hexapoda (12)]. Аргументом в пользу того, что диандрический аппарат является исходным для современных уховерток, может быть тот факт, что из двух групп уховерток, у которых в личиночном состоянии сохраняются многочлениковые церки, у Diplatys/fg пенис диандрический, а у Karschiella/fg пенис похож на диандрический, но лишь с одним асимметрично расположенным телопенисом, т.е. происходит из диандрического (см. Neodermaptera: «Классификации» V). Другим аргументом в пользу первичности диандрического пениса принято считать то, что у некоторых уховерток, самцы которых имеют диандрический пенис, самки имеют вестигии яйцеклада [см. (8b)].
В прошлом считали, что строение гениталий Diploglossata доказывает первичную парность гениталий у самцов уховерток: таксон Diploglossata противопоставляли всем прочим уховерткам (см. ниже, Neodermaptera: «Классификации» I), и при этом было распространено ошибочное утверждение, что внутри непарного пениса Diploglossata проходит пара семенных протоков, открывающихся на вершине пениса парой отверстий; из этого делали вывод, что в ходе эволюции уховерток исходный парный пенис разными способами преобразовывался в непарный – либо путем слияния пары пенисов в один (у Diploglossata), либо путем редукции левого пениса (у Monandria). Однако на самом деле у Diploglossata пенис монандрический: за парные протоки ошибочно принимали пару слепозаканчивающихся впячивающихся мешочков, тогда как истинный семенной проток непарный и открывается на вершине пениса между отверстиями мешочков (Davies 1966b).
Другим аргументом в пользу исходной парности гениталий уховерток считали утверждение о том, что парный пенис обнаружен у одного из представителей Archidermaptera; в данном случае за пенис самца был принят яйцеклад самки [см. ниже, Archidermaptera (4)].
Детали строения гениталий достоверно известны только для Neodermaptera, но не для вымерших групп; известно лишь, что некоторые Archidermaptera имеют подвижные парамеры [см. Archidermaptera (4)]. Так что неясно, к какому таксону относятся описанные выше особенности гениталий Dermatoptera – к Dermapteroidea, Brachydermaptera или Neodermaptera.
(10) Модификация церков. Мышцы, идущие от оснований церков к X тергиту брюшка [см. Amyocerata (8)], направлены не только вперед, но и назад: от основания каждого церка, помимо идущих вперед латеральных мышц-раздвигателей и медиальных мышц-сдвигателей (имеющихся у многих насекомых), отходит медиальная каудальная мышца-раздвигатель; она отходит от медиального края основания церка, идет назад-медиально и крепится к заднему выступу тергита, который простирается позади оснований церков. Латеральные мышцы, исходно служащие раздвигателями церков, у уховерток могут крепиться к основаниям церков медиальнее их мыщелков, и в этом случае они служат не раздвигателями, а сдвигателями церков; в таком случае функцию раздвигания церков выполняют только медиально-каудальные мышцы. У личинок X тергит брюшка цельный, а медиальные каудальные мышцы церков крепятся к его задней части (рис. 8.7.3A). У имаго большинства Neodermaptera задний выступ тергита, к которому крепятся медиально-каудальные раздвигатели церков, преобразован в так называемый пигидий (pygidium): он отчленен от остальной части тергита, загнут вентрально и имеет вид подвижного придатка, выступающего назад между церками; медиально-каудальные мышцы идут от оснований церков назад и крепятся внутри пигидия (рис. 8.7.3B).
У Paradermaptera (имеющих клещевидные церки) и у Diploglossata (имеющих прямые одночлениковые церки) пигидий не отчленен от остальной части тергита, так что вместо подвижного пигидия имеется сквамопигидий (squamopygidium) – неподвижный выступ X тергита брюшка. Это либо результат вторичного срастания, либо исходное состояние; во втором случае отчленение пигидия от тергита является аутапоморфией таксона Eudermaptera (см. ниже, Neodermaptera: «Классификации» II).
Некоторые авторы считают, что пигидий уховерток гомологичен эпипрокту; в действительности это не так, поскольку к эпипрокту мышцы от оснований церков никогда не идут; истинный эпипрокт находится между пигидием и анальным отверстием. Удлиненный пигидий внешне несколько напоминает парацерк; однако парацерк здесь отсутствует [см. Metapterygota (4)]; в отличие от пигидия, внутри парацерка, как и вообще внутри каудалий, мышцы никогда не крепятся.
В остальном десятый уромер сохраняет примитивное строение: его тергит на вентральной стороне не замкнут, так что парапрокты причленяются к IX уромеру; это положение сохраняется у всех уховерток, несмотря на то, что у подавляющего большинства современных уховерток в связи с преобразованием церков в мощные хватательные клещи, X тергит брюшка очень сильно увеличен. Церки всегда развиты, бывают многочлениковыми или одночлениковыми, неспециализированы или специализированы как клещи [см. ниже, Protelytroptera (3), Archidermaptera (5) и Neodermaptera (2)].
Признаки Dermatoptera неясного филогенетического значения
(11) Восьмичлениковая антенна личинки. Флагеллум антенны [см. Amyocerata (1)] нитевидный или четководный (рис. 8.7.1), у личинки первого возраста имеет постоянное число члеников, равное 6; т.е. антенна в целом 8-члениковая; при последующих линьках число члеников увеличивается за счет разделения только первого членика флагеллума (в отличие от некоторых других Amyocerata, у которых разделяются несколько проксимальных члеников флагеллума). У имаго число члеников флагеллума различно: в разных таксонах от 6 до примерно 50, иногда варьирует индивидуально [несмотря на постоянное число личиночных возрастов – см. Pterygota (4Ca)]. Развитие антенны изучено для современных уховерток (Giles 1961, Davies 1966a); но для вымерших таксонов Protelytroptera и Archidermaptera этот признак неизвестен.
(12) Политрофический яичник. Яичник мероистический политрофический [см. Hexapoda (12)]. В каждом фолликуле находится один полиплоидный трофоцит и одна яйцеклетка. Такое строение яичников описано для разных современных уховерток, включая Diploglossata и Plecodermaptera (Bonhag 1958). У разных уховерток такое строение возникает разными путями: у одних видов сначала при делении одного цистобласта образуется большой кластер из многих яйцеклеток и трофоцитов, а затем этот кластер разделяется на фрагменты, каждый из которых содержит одну яйцеклетку и один или несколько трофоцитов; у других видов каждый цистобласт делится всего один раз, образуя одну яйцеклетку и один трофоцит (Büning 1996). Помимо Dermatoptera, политрофические яичники имеются у Panpsocoptera и многих Metabola (см. Указатель признаков [4.11.4]), но у этих насекомых в каждом фолликуле обычно имеется не один, а несколько трофоцитов. Вероятно, политрофические яичники возникали и исчезали несколько раз, но было высказано предположение, что их наличие является синапоморфией Dermatoptera, Acercaria и Metabola (см. «Систематическое положение»).
(13) Хромосомы голокинетические [см. Eucaryota (2c)]. Этот признак изучен для нескольких видов, включая представителя Diploglossata (White 1976). Неуникальная апоморфия (см. Указатель признаков [1.12.1]).
Плезиоморфии и изменчивые признаки Dermatoptera
(14) Число тарсомеров. Лапки исходно пятичлениковые [см. Amyocerata (5)]. У всех современных представителей лапки 3-члениковые (рис. 8.7.1A) [см. ниже, Neodermaptera (1)], но у примитивных вымерших представителей лапки достоверно сохраняют все 5 члеников (рис. 8.7.2B–C) [см. ниже, Archidermaptera (1)].
Геологический возраст. См. отдельно Dermapteroidea, Brachydermaptera и Neodermaptera.
Систематическое положение Dermatoptera. Помимо принятого здесь отнесения Dermatoptera к Tegminoptera, предлагались следующие классификации.
I. Объединение Dermatoptera, Acercaria и Metabola в таксон Meroista Büning 1996 на основании того, что яичники мероистические политрофические [см. Hexapoda (12) и Dermatoptera (12)]. В этом случае считают, что в некоторых группах в составе Acercaria и Metabola яичники вторично стали паноистическими или телотрофическими. В свою очередь предлагалось объединять Meroista и Plecoptera в таксон Promeroista (см. Plecoptera: «Систематическое положение» V). В качестве синапоморфии Dermatoptera, Acercaria и Metabola, помимо строения яичников, называют также разделение стилигеров IX уромера самки на проксимальный и дистальный отделы (Klass 2003); этот признак можно интерпретировать и как синапоморфию Dermatoptera и Eumetabola [см. Eumetabola (1)].
II. Объединение Dermatoptera, Embioptera и Plecoptera в таксон Panplecoptera (см. Embioptera: «Систематическое положение» V).
III. Объединение Zoraptera и Dermatoptera в таксон Haplocercata (см. Zoraptera: «Систематическое положение» V).
VIII-1.2.2.4.B. Классификация Dermatoptera с учетом вымерших групп
Филогения Dermatoptera представляется следующим образом:
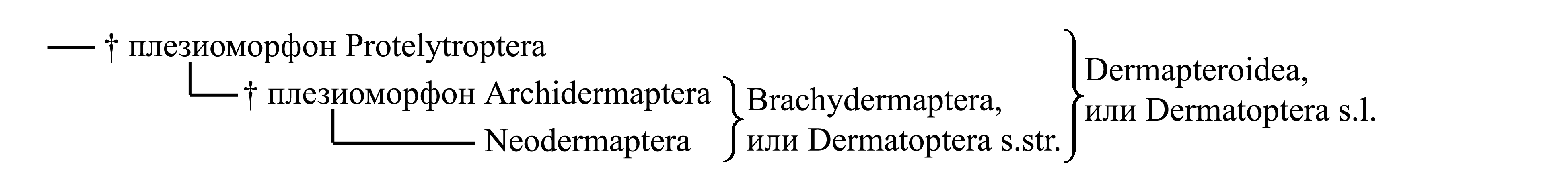
Ниже рассматривается следующая классификация уховерток:
1. Dermapteroidea, или Dermatoptera s.1, или Forficula/f1=g2)
1.1. † Плезиоморфон Protelytroptera, или Protelytron/fg1)
1.2. Brachydermaptera, или Dermatoptera s.2, или Forficula/f2=g3)
1.2.1. † Плезиоморфон Archidermaptera, или Protodiplatys/fg1)
1.2.2. Neodermaptera, или Dermatoptera s.3, или Forficula/f3=g4)
![]() 1. Dermapteroidea, или Dermatoptera s.1,
или Forficula/f1=g3
1. Dermapteroidea, или Dermatoptera s.1,
или Forficula/f1=g3
![]() 1.1. † Плезиоморфон Protelytroptera,
или Protelytron/fg1
1.1. † Плезиоморфон Protelytroptera,
или Protelytron/fg1
![]() 1.2. Brachydermaptera, или Dermatoptera
s.2, или Forficula/f2=g3
1.2. Brachydermaptera, или Dermatoptera
s.2, или Forficula/f2=g3
![]() 1.2.1. † Плезиоморфон Archidermaptera,
или Protodiplatys/fg1
1.2.1. † Плезиоморфон Archidermaptera,
или Protodiplatys/fg1
![]() 1.2.2. Neodermaptera, или Dermatoptera s.3,
или Forficula/f3=g5
1.2.2. Neodermaptera, или Dermatoptera s.3,
или Forficula/f3=g5