КЛАДОЭНДЕЗИС НАСЕКОМЫХ

![]()
|
КЛАДОЭНДЕЗИС НАСЕКОМЫХ |
|||||||
|
|
|
fg | |||||
VIII-1.2.2.1.B-1.2. Cryptovipositoria Kluge 2010, или Pandictyoptera s.str.
(Somobialia![]() Eucaryota
Eucaryota![]() Metazoa
Metazoa![]() Monostomata
Monostomata![]() Haemataria
Haemataria![]() Peritoneata
Peritoneata![]() Metameria
Metameria![]()
Gnathopoda![]() Euarthropoda
Euarthropoda![]() Mandibulata
Mandibulata![]() Atelocerata
Atelocerata![]() Hexapoda
Hexapoda![]() Amyocerata
Amyocerata![]()
Pterygota![]() Metapterygota
Metapterygota![]() Neoptera
Neoptera![]() Rhipineoptera
Rhipineoptera![]() Tegminoptera
Tegminoptera
Pandictyoptera![]() – Holopandictyoptera
– Holopandictyoptera![]() Cryptovipositoria)
Cryptovipositoria)
(рис.
8.4.1C–D, 8.4.2–8.4.6)
(рис.
8.4.1C–D,
8.4.2,
8.4.3,
8.4.4,
8.4.5,
8.4.6)
|
Название, однозначно соответствующее по циркумскрипции: — Cryptovipositoria Kluge 2010a. Названия, неоднозначно соответствующие по циркумскрипции (соответствующие также Mantis/f1=Blatta/fg1) – см. выше, Pandictyoptera. По циркумскрипции также соответствует таксонам под типифицированными названиями, образованными от родового названия Blatta. Иерархическое название: Mantis/f2=Blatta/g2 [f:1802:270; g:L.1758(gen.193)] (sine. Mylacris; incl. Termes). |
|
Ранг. От подотряда до надотряда.
Аутапоморфия, достоверно относимая к Cryptovipositoria
(1) Модификация яйцеклада. У самки половой аппарат модифицирован для формирования оотеки (ootheca) – оболочки, покрывающей пакет яиц. VII стернит брюшка увеличен, его задний край выступает далеко назад (рис. 8.4.2D, 8.4.3A, 8.4.5); эта выступающая назад лопасть, называемая гипопигий (hypopygium), или генитальная пластинка, закрывает вентральные стороны VIII и IX сегментов брюшка, образуя внешнюю генитальную камеру, или вестибулюм (vestibulum): вентральной стенкой вестибулюма служит дорсальная мембранозная кутикула гипопигия, а дорсальной стенкой – VIII и IX стерниты брюшка. Яйцеклад [образованный придатками VIII и IX сегментов брюшка – см. Amyocerata (7)] крепится внутри вестибулюма, укорочен и модифицирован следующим образом. Три пары створок, образующих яйцеклад, сцеплены друг с другом с каждой стороны: створки первой и второй пары (VIII и IX кинетапофизы) сохраняют прочную исходную сцепку посредством олистетера, а створка третьей пары (IX стилигер) непрочно сцеплена с ними благодаря комплементарным выпуклостям и вогнутостям той ли иной формы на обращенных друг к другу сторонах створок (рис. 8.4.2E, 8.4.5A–C). При этом створки каждой пары разделены почти от самого основания, так что яйцеклад состоит из левой и правой половин. Кутикула створок в значительной части эластичная. Под действием мышц, идущих к каждой створке [см. Amyocerata (7)], три сцепленные друг с другом створки каждой половины яйцеклада способны деформироваться, благодаря чему яйцеклад способен совершать движения, обеспечивающие постройку оотеки.
В отличие от кутикулы насекомых, оболочка оотеки не содержит хитин и состоит из задубленных белков, часто с кристаллами оксалата кальция. Вещество оотеки образуется из секрета пары придаточных коллетериальных желез [см. Hexapoda (12)]; эти железы имеют ветвистую форму и открываются парой протоков между створками яйцеклада. Левая коллетериальная железа имеет больший размер; ее секрет содержит структурный белок и вещества, служащие предшественниками при синтезе хинона. Секрет правой коллетериальной железы содержит глюкозидазу. При излиянии секрета обеих желез в вестибулюм глюкозидаза из правой железы реагирует с веществами из левой железы, в результате чего образуется хинон; хинон задубливает белок, входящий в состав секрета левой железы, в результате чего образуется твердая оболочка оотеки (Roth 1968).
У богомолов вершина яйцеклада выступает из вестибулюма и формирует снаружи от него крупную оотеку, прикрепленную к субстрату [см. ниже, Raptoriae (4)]. У тараканов яйцеклад полностью скрыт в вестибулюме и участвует в формировании оотеки внутри вестибулюма [см. ниже, Oothecophora (2)]. У термитов способность образовывать оотеку вторично утрачена [см. ниже, Isoptera (6)].
У самок некоторых видов на вершинах третьих створок яйцеклада (т.е. IX унистилигеров) сохраняются стерностили [без мышц – см. Metapterygota (5)]; иногда стерностили отсутствуют у имаго, но имеются у личинок [в отличие от самца, у которого стерностили сохраняются и у имаго – см. Pandictyoptera (8)].
Геологический возраст. Ранний мел – ныне (рис. 2.2); возможен более ранний возраст (см. ниже, Neoblattariae).
Классификации Cryptovipositoria.
I. Деление Cryptovipositoria на Oothecaria и Isoptera. Таксон Oothecaria Verhoeff 1902b {synn.circ.: Homoneura Enderlein 1903 (интерпретация названия Nomoneura Brauer 1885) = Palaeopteradelphia Crampton 1915 = Blattoptera Schwanwitsch 1946; по циркумскрипции также соответствует: «Dictyoptera» sensu Enderlein 1904 (non Dictuoptera Leach 1815a – см. выше, комментарий к Pandictyoptera)} объединяет тараканов (Neoblattariae) и богомолов (Raptoriae). Oothecaria отличаются от Isoptera лишь симплезиоморфиями: яйца откладываются в оотеку [см. (1)] (тогда как у Isoptera оотека вестигиальная или утрачена), гениталии самца имеют три фалломера [см. Pandictyoptera (8)] (тогда как у Isoptera фалломеры редуцированы). Судя по всему, таксон Oothecaria является парафилетическим (см. II).
II. Деление Cryptovipositoria на Raptoriae и Oothecophora. В этом случае богомолы (Raptoriae) противопоставляются голофилетическрому таксону Oothecophora, куда относятся тараканы (Neonlattariae) и термиты (Isoptera).
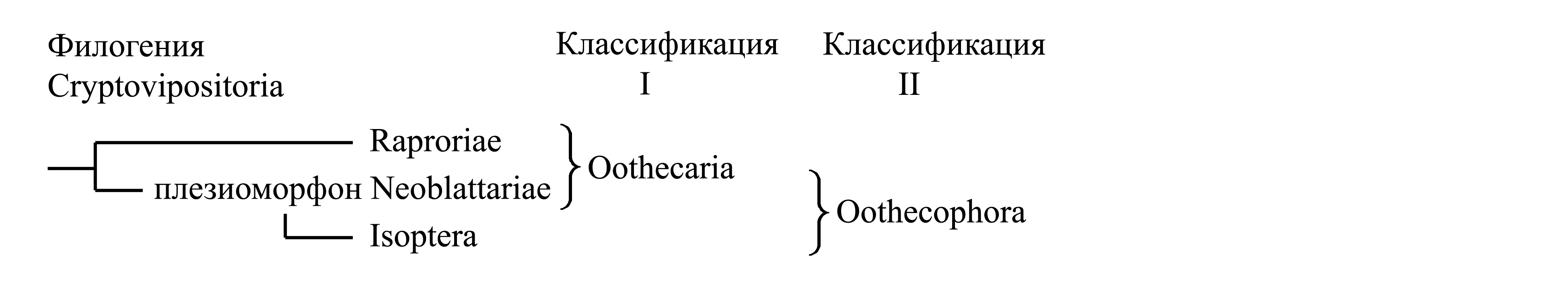
Здесь мы принимаем деление Cryptovipositoria на Raptoriae и Oothecophora (с. 0000).
Cryptovipositoria, или Pandictyoptera s.str., или Mantis/f2=Blatta/g2
![]() 1. Raptoriae, или
Mantis/f3=g1
– богомолы
1. Raptoriae, или
Mantis/f3=g1
– богомолы
![]() 2. Oothecophora, или
Termes/f1=Blatta/g3
2. Oothecophora, или
Termes/f1=Blatta/g3
![]() 2.1. Плезиоморфон Neoblattariae,
или Blatta/f1=g4 – настоящие тараканы
2.1. Плезиоморфон Neoblattariae,
или Blatta/f1=g4 – настоящие тараканы
![]() 2.2. Isoptera, или
Termes/f2=g1
– термиты
2.2. Isoptera, или
Termes/f2=g1
– термиты