КЛАДОЭНДЕЗИС НАСЕКОМЫХ

![]()
|
КЛАДОЭНДЕЗИС НАСЕКОМЫХ |
|||||||
|
|
|
fg | |||||
VIII-1.2.2.1. Pandictyoptera Crampton 1917
(Somobialia![]() Eucaryota
Eucaryota![]() Metazoa
Metazoa![]() Monostomata
Monostomata![]() Haemataria
Haemataria![]() Peritoneata
Peritoneata![]() Metameria
Metameria![]()
Gnathopoda![]() Euarthropoda
Euarthropoda![]() Mandibulata
Mandibulata![]() Atelocerata
Atelocerata![]() Hexapoda
Hexapoda![]() Amyocerata
Amyocerata![]()
Pterygota![]() Metapterygota
Metapterygota![]() Neoptera
Neoptera![]() Rhipineoptera
Rhipineoptera![]() Tegminoptera
Tegminoptera![]() Pandictyoptera)
Pandictyoptera)
(рис. 8.4.1–8.4.6)
(рис. 8.4.1,
8.4.2, 8.4.3,
8.4.4, 8.4.5,
8.4.6)
|
Названия, соответствующие по циркумскрипции и Blatta/g1, и Blatta/g2: — Pandictyoptera Crampton 1917; — Panisoptera Crampton 1919; — Blattopteroidea Martynov 1924a; — Isopteria Crampton 1938; — Dictuopteroida Kevan 1976; — Oothecariformia Zompro 2005. По циркумскрипции также соответствует: — «Dictyoptera» sensu Bolivar 1898 (исходно Dictiopteros, в испанском написании) (non Dictyoptera Clairville 1798, nec Dictyoptera Brullé 1832, nec Dictyoptera Dohrn 1866); — «Dictuoptera» sensu Vickery & Kevan 1983 (non Leach 1815a). — таксонам под типифицированными названиями, образованными от родового названия Blatta – Blattidea и др. Иерархическое название: Mantis/f1–2=Blatta/g1–2 [f:1802:270; g:L.1758(gen.193)] (incl. Mylacris, Termes). |
|
Ранг. Обычно отряд или надотряд.
Название. В литературе этот таксон, объединяющий богомолов, тараканов и термитов, часто называют «Dictyoptera» или «Dictuoptera», приписывая обоим названиям авторство «Leach 1815». В действительности, в статье Лича (Leach 1815a) под названием Dictuoptera описан отряд, включающий только род Blatta, т.е. соответствующий Neoblattariae (см. ниже), тогда как богомолы и термиты отнесены к другим отрядам. В последующих публикациях того же автора название «Dictuoptera» использовалось в том же написании и в том же значении. Название Dictyoptera (сетчатокрылые) независимо давалось для разных групп насекомых: Dictyoptera Clairville 1798 является синонимом Neuroptera Linnaeus 1758 (см. Pterygota: «Классификации»); Dictyoptera Brullé 1832 – таксон, объединяющий амфибиотических насекомых с неполным превращением, т.е. стрекоз, поденок и веснянок (см. Ephemeroptera: «Систематическое положение» III); Dictyoptera Dohrn 1866 является объективным синонимом названия Palaeodictyoptera Goldenberg 1877 и относится к вымершей группе насекомых (см. Protorrhynchota: «Классификация»). Кроме того, название Dictyoptera употреблялось в том же значении, что Oothecaria (см. ниже, Cryptovipositoria: «Классификации» I) и в том же значении, что Raptoriae (см. ниже).
Если рассматривать только современных представителей Pandictyoptera, то таксон Pandictyoptera выглядит четко очерченным. В этом случае все названия, перечисленные выше, однозначно соответствуют по циркумскрипции. Однако существуют вымершие палеозойские и мезозойские тараканы, которые имеют явное родство с настоящими тараканами, но не укладываются в таксон Pandictyoptera, охарактеризованный только по строению современных представителей. Эти формы вместе с современными тараканами, термитами и богомолами образуют голофилетический таксон Pandictyoptera s.1., или Holopandictyoptera (или Mantis/f1=Blatta/g1), внутри которой находится подчиненный таксон Pandictyoptera s.str., или Cryptovipositoria (или Mantis/f2=Blatta/g2). Мы не имеем возможности рассмотреть все интересующие нас признаки этих соподчиненных таксонов, так как для вымерших форм известны лишь немногие детали строения. Поэтому здесь характеристика пандиктиоптер разбита на две части. В первой части – «А. Общая характеристика Pandictyoptera» рассматриваются признаки, про многие из которых неясно, относятся ли они ко всем Pandictyoptera в широком смысле, или лишь к Pandictyoptera в узком смысле. Во второй части – «B. Классификация Pandictyoptera с учетом вымерших групп» рассматриваются признаки, распределение которых в этих группах достоверно известно.
VIII-1.2.2.1.A. Общая характеристика Pandictyoptera
Аутапоморфии Pandictyoptera
(1) Перфорированный тенториум. Тенториум усложненного строения, т.н. перфорированный (рис. 8.4.1C): передние руки тенториума впереди от их мест слияния с задним тенториальным мостом [т.е. впереди корпотенториума – см. Pterygota (1)] расширены и слиты друг с другом, образуя непарную центральную пластинку; в месте их слияния сохраняется медиальный продольный шов; между центральной пластинкой и корпотенториумом сохраняется непарное тенториальное отверстие. Центральная пластинка находится позади (вентральнее) пищевода, но впереди (дорсальнее) окологлоточных коннектив [тогда как корпотенториум находится позади (вентральнее) пищевода и окологлоточных коннектив], так что окологлоточные коннективы (соединяющие надглоточный ганглий с подглоточным) проходят сквозь тенториальное отверстие.
Наличие центральной пластинки и тенториального отверстия свойственно всем Raptoriae, Neoblattariae и Isoptera и является уникальной апоморфией Pandictyoptera. Для вымерших Palaeoblattariae строение тенториума неизвестно, так что неясно, является ли это аутапоморфией Holopandictyoptera в целом или лишь аутапоморфией Cryptovipositoria.
(2) Модификация шейных склеритов. Голова очень подвижная благодаря тому, что цервикальные склериты [см. Pterygota (2)] имеют характерное строение (рис. 8.4.1D). Каждый из пары цервикальных склеритов разделен на две подвижно сочлененные части – передний цервикальный склерит (сочлененный с головой) и задний цервикальный склерит (сочлененный с переднегрудью); задний цервикальный склерит сильно расширен и выдается вентрально (медиально), так что на вентральной стороне шеи левый и правый задние цервикальные склериты сильно сближены или соприкасаются; у некоторых они Raptoriae слиты. Такое строение цервикальных склеритов достоверно известно для современных Raptoriae, Neoblattariae и Isoptera. Для вымерших Palaeoblattariae строение цервикальных склеритов неизвестно, так что не вполне ясно, является ли это аутапоморфией Holopandictyoptera в целом или лишь аутапоморфией Cryptovipositoria сходство в общей форме переднегруди и головы у Palaeoblattariae и Neoblattariae позволяет предположить, что их цервикальные склериты также сходны.
Такое строение шейных склеритов обеспечивает большую подвижность головы. У тараканов голова в покое может занимать гипогнатное или опистогнатное положение, а при питании поворачиваться прогнатно (рис. 8.4.1D). У богомолов голова особенно подвижна, что позволяет им охотиться из засады и прицеливаться к добыче, сохраняя неподвижность туловища [см. ниже, Raptoriae (1)].
Сходная конструкция цервикальных склеритов независимо возникла у Amphiesmenoptera, но там задние цервикальные склериты сочленяются с помощью узких вентральных отростков.
(3) Форма пронотума. Пронотум исходно широкий, овальный, с распластанными краями, так что его передний край налегает на голову, задний край налегает на основание крыльев, а боковые края распластаны в стороны в виде широких закругленных лопастей – паранотумов. Такой широкоовальный пронотум характерен для тараканов – Palaeoblattariae и Neoblattariae (рис. 8.4.1A–B, D, 8.4.3A). У термитов (Isoptera) пронотум уменьшен, но сохраняет небольшие распластанные края, налегая передним краем на задний край головы (рис. 8.4.6D). У различных Raptoriae пронотум разнообразной формы: у некоторых видов он широкой, с большими распластанными краями, как у тараканов; у видов с удлиненной и суженной переднегрудью [см. ниже, Raptoriae (1)] распластанные края пронотума могут быть менее выражены.
(4) Модификация птероторакса. Птероторакс имеет характерное строение (рис. 8.4.3A). На среднегруди и заднегруди постнотум и фрагмы редуцированы, продольные спинные мышцы сильно утоньшены. В то время как у других Neoptera нотум (т.е. передняя часть тергита) и постнотум (т.е. задняя часть тергита) способны сгибаться относительно друг друга под действием мощных продольных спинных мышц, крепящихся к фрагмам [см. Pterygota (4Abβ)], у Pandictyoptera этот механизм не развит, и полет осуществляется за счет действия крыловых мышц прямого действия. Плейриты птероторакса более или менее сильно наклонены [дорсальной стороной вперед – см. Neoptera (2)]. У всех трех пар ног тазики сильно увеличены, длинные, направлены назад и сближены между собой. Субалярно-коксальная мышца увеличена и служит опускателем крыла прямого действия [см. Pterygota (4Abγ)]; в связи с этим у летающих форм задний отдел среднего и заднего тазика, вмещающий основание субалярно-коксальной мышцы, утолщен и отделен швом от остальной части тазика. Стерниты среднегруди и заднегруди сужены; стернальные аподемы [см. Pterygota (3)] иногда более или менее расставлены (как у большинства Rhipineoptera), а иногда расходятся из одной точки (что напоминает криптостернию многих Eumetabola). В связи с сильным наклоном заднегруди первый стернит брюшка редуцирован (то же у многих других насекомых – см. Указатель признаков [3.9]).
Такое строение птероторакса достоверно известно для современных Raptoriae, Neoblattariae и Isoptera. Для вымерших Palaeoblattariae детали строения птероторакса неизвестны, так что не вполне ясно, являются ли названные признаки аутапоморфиями всех Holopandictyoptera, или это аутапоморфии лишь Cryptovipositoria; сходство в общей форме тела у Palaeoblattariae и Neoblattariae позволяет предположить, что детали строения птероторакса у них также сходны.
(5) Дополнительные шпоры на ногах. Помимо двух апикальных гиподермосодержащих шпор голени, на ногах имеются дополнительные гиподермосодержащие шпоры – на голени (5a) и на бедре (5b). Кутикула шпор подостлана многоклеточной гиподермой, так что перед линькой новая шпора закладывается внутри старой, однако внешне шпоры напоминают щетинки [производные оболочечных клеток нейрона – см. Euarthropoda (5a)] и могут быть легко с ними спутаны. В случаях, когда имеются и первичные, и дополнительные шпоры, у личинки первого возраста имеются только две первичные апикальные шпоры голени и одна дополнительная апикальная шпора бедра, тогда как на месте других дополнительных шпор находятся щетинки. При последующей линьке на месте этих щетинок возникают шпоры (рис. 8.4.4).
(5 a) Шпоры голени. На голени, помимо двух первичных вершинных гиподермосодержащих шпор [см. Neoptera (4)], могут быть развиты дополнительные гиподермосодержащие шпоры, расположенные не только у вершины, а по всей голени. У Palaeoblattariae и Neoblattariae дополнительные шпоры беспорядочно разбросаны со всех сторон голени (рис. 8.4.4B) (что, вероятно, исходно для Pandictyoptera); на хватательных передних ногах богомолов имеется два правильных ряда неподвижных шиповидных шпор на вентральной стороне голени [см. ниже, Raptoriae (1)] (рис. 8.4.2C).
Иногда дополнительные и первичные вершинные шпоры голени утрачены: на средних и задних ногах Raptoriae и на всех ногах Isoptera дополнительные шпоры голени обычно утрачены и имеются лишь две первичные вершинные шпоры; иногда утрачены также и первичные вершинные шпоры.
(5 b) Шпоры бедра. На бедре могут быть развиты следующие гиподермосодержащие шпоры: коленная шпора, шпоры вентрально-переднего ряда и шпоры вентрально-заднего ряда (рис. 8.4.3A, 8.4.4).
Коленная шпора находится на вершине бедра, на его дорсально-передней стороне. На передней ноге коленная шпора находится ближе к вентральному (внутреннему) краю бедра, так что кажется принадлежащей вентрально-переднему ряду шпор. На средней и задней ноге коленная шпора находится ближе к дорсальному (наружному) краю бедра, так что отчетливо удалена от вентрально-переднего ряда шпор. Коленная шпора может быть развита начиная с первого личиночного возраста, когда на месте других шпор бедра имеются лишь щетинки (рис. 8.4.4A). Коленная шпора имеется у многих Palaeoblattariae и Neoblattariae (имеющих, кроме того, многочисленные шпоры на внутренней стороне бедра и на голени) и у многих Raptoriae.
Вентрально-передний и вентрально-задний ряды шпор – это два правильных продольных ряда шпор, обрамляющих вентральную (внутреннюю) сторону бедра. Когда нога согнута в колене, голень прижимается к бедру между этими двумя рядами шпор. Вентрально-передний и вентрально-задний ряды шпор развиты на всех ногах у многих тараканов (как Palaeoblattariae, так и Neoblattariae) и всегда развиты на хватательных передних ногах богомолов [см. ниже, Raptoriae (1)].
Иногда дополнительные шпоры бедра утрачены. У Raptoriae на средних и задних ногах нет двух вентральных рядов шпор, имеются лишь коленные шпоры; иногда коленные шпоры также утрачены. У некоторых Neoblattariae на ногах определенных пар или на всех ногах утрачены шпоры двух вентральных рядов и/или коленные шпоры. У Isoptera все шпоры бедра утрачены.
(6) Модификация преклавуса переднего крыла. Переднее крыло [лишенное краевой жилки и служащее надкрыльем – см. Tegminoptera (1) и (2)] исходно широкое, с выпуклым передним краем, декостализовано: оканчивающиеся на переднем крае жилки Sc и RA короткие, а RS образует много ветвей, также оканчивающихся на переднем крае крыла (рис. 8.4.3A). Такая декостализация переднего крыла характерна для большинства Palaeoblattariae и Neoblattariae; у некоторых тараканов крылья сильно видоизменены и вторично костализованы; у богомолов переднее крыло вторично костализовано благодаря удлинению Sc и RA [см. ниже, Raptoriae (3)] (рис. 8.4.2A); у многих термитов крылья костализованы благодаря слиянию С, Sc и RA и утрате ветвления RS [см. ниже, Isoptera (5)]. У многих Pandictyoptera крылья вестигиальные или утрачены [см. Neoptera (1e)].
(7) Модификация клавуса переднего крыла. На переднем крыле клавус [см. Neoptera (1b)] имеет характерное строение: клавальный желоб имеет форму дуги с выпуклостью вперед, а многочисленные жилки клавуса изогнуты дугообразно так, что их вершины сближены или сливаются (рис. 8.4.2A, 8.4.3A). Такой «тараканий клавус» имеется у многих Palaeoblattariae, Neoblattariae и Raptoriae, но у некоторых видов он вторично редуцирован или утратил характерный облик; у Isoptera он сильно уменьшен и утратил жилкование (рис. 8.4.6A, D). У многих Pandictyoptera крылья вестигиальные или утрачены [см. Neoptera (1e)].
(8) Фалломеры самца. У самца фаллический комплекс [см. Amyocerata (6)] состоит из трех выростов – фалломеров (phallomeri, или phallomera), два из которых находятся по бокам от генитального отверстия (левый и правый фалломеры), а третий – вентральнее генитального отверстия (вентральный фалломер). Половое отверстие может находиться между основаниями этих фалломеров или смещено на один из фалломеров, который в этом случае называется пенисом. Форма фалломеров и их склеротизованных и мембранозных участков специфична для определенных таксонов. Сложная система мышц, начинающихся в IX сегменте, входит в основания фалломеров; кроме того, имеются многочисленные мышцы внутри фалломеров, входящие в различные их части. По форме, склеротизации и мускулатуре все три фалломера различаются – т.е. гениталии ассиметричные; в типичном случае правый фалломер расположен дорсальнее других. Иногда какой-либо из склеритов преобразован в титиллятор – длинный шип или крюк. В определенных таксонах богомолов и тараканов асимметричные гениталии имеют план строения, соответствующий зеркальному отражению исходного.
С вентральной стороны фаллический комплекс прикрыт гипандрием, к заднему краю которого причленены стерностили [без мышц – см. Metapterygota (5)]; у некоторых Neoblattariae и некоторых Isoptera стерностили утрачены.
Такой генитальный аппарат с тремя фалломерами имеется у Neoblattariae и Raptoriae, но редуцирован у Isoptera (Snodgrass 1937). Строение гениталий вымерших Palaeoblattariae неизвестно, так что неясно, относятся ли названные признаки к Pandictyoptera в целом или только к Cryptovipositoria. Помимо Pandictyoptera, асимметричный копулятивный аппарат, состоящий из отдельных фалломеров с половым отверстием в основании, имеется также у Notoptera и некоторых других (в отличие от Saltatoria, Dermatoptera и др., имеющих трубчатый пенис).
(9) Модификация яйцеклада. У самки яйцеклад особым образом модифицирован и целиком или частично скрыт во внешней генитальной камере, образованной увеличенным VII стернитом брюшка; благодаря этому откладываемые яйца оказываются заключенными в оболочку – оотеку [см. ниже, Cryptovipositoria (1)]. Эта особенность свойственна только Pandictyoptera в узком смысле слова (т.е. Cryptovipositoria), тогда как у вымерших Palaeoblattariae сохранялся длинный яйцеклад и, по-видимому, не было внешней генитальной камеры и способности образовывать оотеку [см. ниже, Palaeoblattariae (1)] (рис. 8.4.1A–B). Строение оотеки различается у богомолов [см. ниже, Raptoriae (4)] и тараканов [см. ниже, Oothecophora (2)]; у термитов способность образовывать оотеку утрачена [см. ниже, Isoptera (6)].
Плезиоморфии и изменчивые признаки Pandictyoptera
(10) Церки. Церки [только парные – см. Metapterygota (4)] обычно развиты, многочлениковые (рис. 8.4.2D, 8.4.3A); у некоторых богомолов, тараканов и у многих термитов церки уменьшены, нерасчлененные; у некоторых термитов церки совсем утрачены (например, у солдат Heterotermes/fg).
Геологический возраст. Карбон – ныне (рис. 2.2) (см. ниже, Palaeoblattariae).
VIII-1.2.2.1.B. Классификация Pandictyoptera с учетом вымерших групп
Филогения Pandictyoptera s.l. (или Holopandictyoptera) представляется следующим образом:
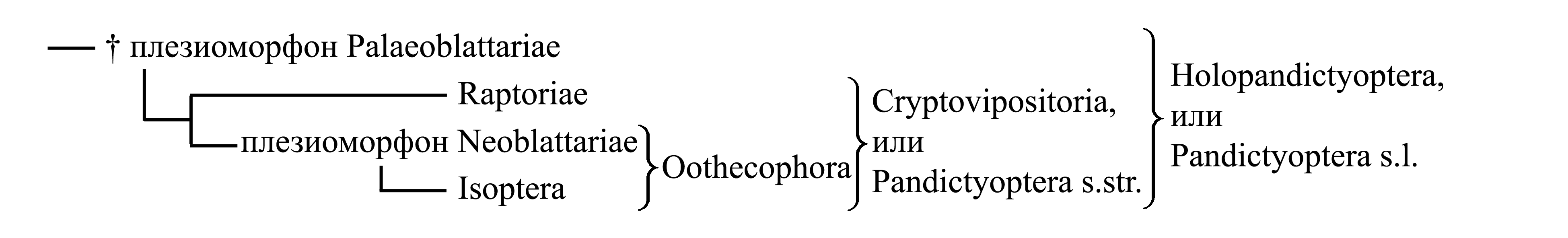
Ниже рассматривается следующая классификация Pandictyoptera:
1. Holopandictyoptera, или Pandictyoptera s.l., или Mantis/f1=Blatta/g1
1.1. †Плезиоморфон Palaeoblattariae, или Mylacris/fg1 – древние тараканы
1.2. Cryptovipositoria, или Pandictyoptera s.str., или Mantis/f2=Blatta/g2
1.2.1. Raptoriae, или Mantis/f3=g1 – богомолы
1.2.2. Oothecophora, или Termes/f1=Blatta/g3
1.2.2.1. Плезиоморфон Neoblattariae, или Blatta/f1=g4 – настоящие тараканы
1.2.2.2. Isoptera, или Termes/f2=g1 – термиты
![]() 1. Holopandictyoptera, или Pandictyoptera
s.l., или Mantis/f1=Blatta/g1
1. Holopandictyoptera, или Pandictyoptera
s.l., или Mantis/f1=Blatta/g1
![]() 1.1. †Плезиоморфон Palaeoblattariae,
или Mylacris/fg1 – древние тараканы
1.1. †Плезиоморфон Palaeoblattariae,
или Mylacris/fg1 – древние тараканы
![]() 1.2. Cryptovipositoria, или Pandictyoptera
s.str., или Mantis/f2=Blatta/g2
1.2. Cryptovipositoria, или Pandictyoptera
s.str., или Mantis/f2=Blatta/g2
![]() 1.2.1. Raptoriae,
или Mantis/f3=g1
– богомолы
1.2.1. Raptoriae,
или Mantis/f3=g1
– богомолы
![]() 1.2.2. Oothecophora,
или Termes/f1=Blatta/g3
1.2.2. Oothecophora,
или Termes/f1=Blatta/g3
![]() 1.2.2.1. Плезиоморфон Neoblattariae,
или Blatta/f1=g4 – настоящие тараканы
1.2.2.1. Плезиоморфон Neoblattariae,
или Blatta/f1=g4 – настоящие тараканы
![]() 1.2.2.2. Isoptera,
или Termes/f2=g1
– термиты
1.2.2.2. Isoptera,
или Termes/f2=g1
– термиты