КЛАДОЭНДЕЗИС НАСЕКОМЫХ

![]()
|
КЛАДОЭНДЕЗИС НАСЕКОМЫХ |
|||||||
|
|
|
fg | |||||
IX-1.2.2.2.1.2.2. Heteroptera Latreille 1810 – полужесткокрылые, или клопы
(Somobialia![]() Eucaryota
Eucaryota![]() Metazoa
Metazoa![]() Monostomata
Monostomata![]() Haemataria
Haemataria![]() Peritoneata
Peritoneata![]() Metameria
Metameria![]()
Gnathopoda![]() Euarthropoda
Euarthropoda![]() Mandibulata
Mandibulata![]() Atelocerata
Atelocerata![]() Hexapoda
Hexapoda![]() Amyocerata
Amyocerata![]()
Pterygota![]() Metapterygota
Metapterygota![]() Neoptera
Neoptera![]() Eumetabola
Eumetabola![]() Parametabola
Parametabola![]() Acercaria
Acercaria![]() Condylognatha
Condylognatha![]()
Arthroidignatha![]() Hemelytrata
Hemelytrata![]() Heteropteroidea
Heteropteroidea![]() Heteroptera)
Heteroptera)
(рис. 9.8.2)
|
Названия, неоднозначно соответствующие по циркумскрипции (формально соответствующие также Heteropteroidea): — Coleopterohymenoptera Schaeffer 1766; — Dermaptera Retzius 1783 (non Dermaptera De Geer 1773); — Heteroptera Latreille 1810; — Styrelytra Rafinesque 1815; — Heterata Billberg 1820; — Frontirostria Zetterstedt 1828 (non Frontirostres Dumeril 1805); — Coridaria Haeckel 1896; — Heteropterida Pearce 1936. По циркумскрипции также соответствует: — «Hemiptera» sensu Leach 1815a (non Linnaeus 1758); — «Frontirostres» sensu Fallén 1814 (non Dumeril 1805). — таксонам под типифицированными названиями, образованными от родового названия Cimex: Cimicida, Cimicina, Cimicaria и др. Иерархическое название: Cimex/f5=Notonecta/g2 [f:1781; g:L.1758(gen.196)] (sine Peloridium; incl. Enicocephalus, Ceratocombus, Gerris, Salda, Pentatoma). |
|
Ранг. Обычно отряд или подотряд.
Название. Часто этому таксону приписывают название «Hemiptera» с авторством Linnaeus 1758; о разных значениях этого названия – см. выше, Arthroidignatha. Названия Frontirostres Dumeril 1805 и Prostomophora Spinola 1850 исходно относились не ко всем, а лишь к части полужесткокрылых.
Аутапоморфии Heteroptera
(1) Модификация головы. На вентральной стороне головы имеется склеротизованная область, расположенная позади внешнего хоботка [видоизмененной нижней губы – см. Arthroidignatha (1d)]; эту область называют либо гулой (gula), либо постгенальным (защечным) мостом (pons postgenalis, или genopons). Таким образом, в отличие от всех прочих Arthroidignatha, у Heteroptera задний край головы на вентральной стороне образован не задним краем основания внешнего хоботка, а задним краем постгенального моста. В результате этого основание внешнего хоботка смещено вперед, и внешний хоботок оказывается отходящим не от задней, а от передней части головы (рис. 9.8.2A). Поскольку тенториум утрачен [см. (2)], неясно, каково исходное расположение постгенального моста относительно задних тенториальных ямок: впереди от них (как у гипостомального моста Mecopteriformia) или позади них (как у гулы некоторых других насекомых – см. Указатель признаков [1.10]).
Задняя часть цельной склеротизованной головной капсулы, включающей постгенальный мост, отчасти вставлена в переднегрудь; несмотря на это, обычно сохраняется нормальное шейное сочленение с парой цервикальных склеритов; лишь у Cephalonotera голова слита с переднегрудью [см. Pterygota (2)].
Орган Эванса утрачен (в отличие от Auchenorrhyncha и Coleorrhyncha) [см. Hemelytrata (5)].
(2) Утрата тенториума. Весь тенториум [см. Pterygota (1)] утрачен. Неуникальная апоморфия; среди Arthroidignatha частичная редукция тенториума встречается также у некоторых Auchenorrhyncha.
(3) Четырехчлениковый хоботок. Внешний хоботок [исходно 3-члениковый – см. Arthroidignatha (1d)] состоит из 4 члеников благодаря тому, что мембрана между головной капсулой [замкнутой вентрально – см. (1)] и основанием внешнего хоботка склеротизована и образует вторичный проксимальный (первый) членик (рис. 9.8.2A). О вторичном происхождении этого проксимального членика свидетельствует то, что от основания следующего за ним второго членика отходит аподема, проходящая сквозь первый членик, тогда как у прочих Arthroidignatha гомологичная ей аподема отходит от основания первого членика [см. Arthroidignatha (1d)]. У некоторых Heteroptera внешний хоботок вторично стал 3-члениковый из-за редукции этого проксимального вторичного членика; у Sandaliorrhyncha внешний хоботок утратил членистость.
(4) Модификация антенн. У большинства клопов флагеллум [см. Amyocerata (1)] состоит из 2 члеников – базифлагеллума (basiflagellum) и дистифлагеллума (distiflagellum); все 4 членика антенны (скапус, педицеллус и оба членика флагеллума), как правило, длинные; между ними находятся еще два маленьких интеркалярных членика: один между педицеллусом и базифлагеллумом, второй между базифлагеллумом и дистифлагеллумом (рис. 9.8.2A). 4 крупных членика антенны (скапус, педицеллус и два членика флагеллума) развиваются в эмбриогенезе, так что неизменны у личинок всех возрастов и у имаго [см. Hemelytrata (4)]; 2 интеркалярных членика обычно развиваются в постэмбриогенезе благодаря склеротизации сочленовной мембраны.
В различных группах клопов имеются отклонения от этой схемы (Zrzavy 1990a, 1992b). В некоторых случаях антенна у имаго выглядит 5-члениковой или с бóльшим числом члеников благодаря вторичному разделению одного из исходных четырех члеников; в этих случаях у личинки антенна имеет нормальное 4-члениковое строение, а дополнительное расчленение возникает только при линьке на имаго. В других случаях, наоборот, число члеников антенны сокращено. У некоторых клопов интеркалярные членики могут быть втянуты внутрь крупных члеников и в этом случае не видны. Иногда интеркалярные членики утрачены. Иногда число крупных члеников меньше четырех (3, 2 или 1) благодаря слиянию между собой тех или иных члеников (скапуса, педицеллуса, базифлагеллума и/или дистифлагеллума). У некоторых Hydrocorisae (имеющих уменьшенные антенны – см. ниже) у молодой личинки антенна нерасчлененная, и ее разделение на 4 или лишь 3 членика возникает в постэмбриональном развитии.
Помимо Heteroptera интеркалярный членик или склеротизованное кольцо между педицеллусом и первым члеником флагеллума встречается также у некоторых других насекомых (у некоторых Thysanoptera, Diptera); наличие двух интеркалярных члеников является уникальной аутапоморфией.
(5) Утрата медиального глазка. Медиальный простой глазок утрачен; неуникальная апоморфия [см. Amyocerata (4)]. Часто имеется пара широко расставленных простых глазков [только у имаго – см. Eumetabola (2)]. Иногда все глазки утрачены. При этом фасеточные глаза обычно развиты, в редких случаях утрачены.
(6) Пахучие железы. Имеются пахучие железы – дорсальные абдоминальные, наиболее развитые у личинок [см. (7a)] и вентральные метаторакальные, развитые у имаго [см. (7b)] (Schuch & Slater 1995). Эти железы в большинстве случаев выделяют ядовитый секрет и выполняют защитную функцию; иногда их секрет выполняет иную функцию – половую, агрегационную или функцию сигнала тревоги.
(6 a) Дорсальные железы. Дорсальные абдоминальные пахучие железы представляют собой впячивания покровов, отходящие от поверхности; они расположены в IV–VII сегментах брюшка (уромерах) или в некоторых их них; на одном уромере может быть одна непарная железа, открывающаяся непарным отверстием, или пара желез с парой отверстий; отверстия желез открываются на переднем крае соответствующего тергита. Функционирующие железы во всех 4 уромерах встречаются редко (у некоторых Trichotelocera), чаще железы имеются лишь в одном, двух или трех из этих сегментов; наиболее обычным является наличие желез в IV–VI уромерах или только в IV уромере. Иногда железы утрачены.
Дорсальные абдоминальные железы обычно развиты у личинок. У имаго (тергиты брюшка которых закрыты сложенными крыльями) дорсальные абдоминальные железы обычно отсутствуют или имеются в виде нефункционирующих вестигиев (рис. 9.8.2A), в редких случаях развиты и функционируют.
(6 b) Вентральные железы. Вентральные пахучие железы представляют собой впячивания покровов, отходящие от стернита заднегруди. Отверстие каждой железы имеет крышечку, открывающуюся специальной мышцей.
У клопов плезиоморфона Archeocorisae (см. «Классификации» IV) непарная или парная железа открывается непарным или парным отверстием на границе стернита заднегруди и первого стернита брюшка (омфалический тип по Carayon 1971).
У прочих клопов (образующих единую филогенетическую ветвь – см. «Классификации» IV и V) непарная или парная железа открывается парным отверстием в основании парной заднегрудной стернальной аподемы [т.е. фурки – см. Pterygota (3)] (диастомический тип по Carayon 1971). Снаружи фуркальная ямка сильно вытянута в дорсо-латеральном направлении, образуя желоб, который обходит основание задней ноги спереди и заканчивается на заднегрудном плейрите (рис. 9.8.2). Иногда желоб замкнут в канал, и тогда секрет железы выделяется наружу из отверстия на его конце; пара таких отверстий на боках тела представляют собой вторичные отверстия пахучих желез. Поверхность кутикулы вокруг желоба или вокруг отверстия канала может иметь ту или иную пористую фактуру и служит испарителем секрета железы.
Вентральные метаторакальные пахучие железы имеются только у имаго; они бывают одинаково развиты у обоих полов, либо лучше развиты у самцов; иногда железы утрачены.
(7) Утрата аролия. Аролий [см. Hemelytrata (6)] утрачен. У личинки и имаго между двумя коготками часто бывает развита пара щетинковидных или иной формы придатков – паремподии (parempodia); иногда имеется не 2, а 3 или 4 паремподия. У Enicocephalus/fg, Ploteres, некоторых Trichotelocera и Hydrocorisae, помимо двух паремподий, имеется щетинка, которую некоторые авторы считают вестигием аролия.
(8) Интеркалярная область заднего крыла. На заднем крыле клавальный желоб [отделяющий преклавальную область от клавуса с жилками Pcu, V1, V2 – см. Hemelytrata (2)] раздвоен V-образно или Y-образно, его ветви расходятся к краю крыла (рис. 9.8.2); иногда между ветвями имеется дополнительная (интеркалярная) продольная жилка.
(9) Смещение дыхалец IV пары. Дыхальца II уромера обычно расположены на вентральной стороне [так что только дыхальца I уромера находятся на дорсальной стороне, а дыхальца II–VIII уромеров находятся на вентральной стороне – см. Hemelytrata (1b)] (рис. 9.8.2A) (в отличие от Auchenorrhyncha и Coleorrhyncha, у которых дыхальца II уромера находятся на дорсальной стороне).
(10) Модификация гениталий самца. В отличие от Coleorrhyncha и Auchenorrhyncha, парамеры [см. Hemelytrata (7)] утратили непосредственное сочленение с пенисом и не имеют мышц, идущих к пенису, так что двигаются лишь мышцами, идущими к ним от стенок генитальной капсулы; основание пениса окружено парой склеритов или цельной склеротизованной рамкой и способно втягиваться в генитальную капсулу (рис. 9.4.2C, 9.8.2A). У многих Heteroptera половой аппарат симметричный, но иногда пенис и парамеры асимметричные, иногда один парамер утрачен; иногда оба парамера утрачены.
Разнообразие Heteroptera. Большинство клопов обитает на суше и питается соком растений [см. Arthroidignatha (1e)]. Ploteres перешли к жизни на поверхности воды, а многие Hydrocorisae обитают под водой (см. «Классификации» I). Некоторые клопы перешли к хищничеству; несколько видов питаются кровью позвоночных.
Размеры. Мелкие или вторично увеличенные [см. Parametabola (9)]: длина тела от 0,5 mm (некоторые Trichotelocera) до 110 мм (некоторые Hydrocorisae).
Геологический возраст. Триас – ныне (рис. 2.2) (Rasnitsyn & Quicke 2002).
Состав и распространение. Около 40 000 видов; всесветно (Schuh & Slater 1995).
Систематическое положение Heteroptera. Помимо принятого здесь отнесения Heteroptera к Heteropteroidea, предлагалось относить их непосредственно к Hemelytrata, либо непосредственно к Arthroidignatha, либо непосредственно к Condylognatha (см. Condylognatha: «Классификации» I; Arthroidignatha: «Классификации» I).
Классификации Heteroptera
I. Деление Heteroptera на Enicocephalus/fg, Trichotelocera, Ploteres, Hydrocorisae, Riparii, Corticolae, Trichophora и Cimex/fg. Бóльшая часть видов клопов распределена между двумя крупными таксонами – Trichophora и Cimex/fg (sine Pentatoma); меньшая часть видов не попадает в эти крупные таксоны, и для них выделяют сравнительно небольшие таксоны того же ранга. Так что всего получаются следующие 8 таксонов равного ранга (часто их считают инфраотрядами, исходя их того, что Heteroptera – подотряд).
1. Enicocephalus/fg [f:1860; g:1837]. Небольшая группа хищных клопов; передние ноги специализированы как хватательные: голень расширена и уплощена к вершине, а лапка укорочена и модифицирована так, что вместе с выступающим медиально-апикальным углом голени образует полуклешню. Передние крылья, если развиты, перепончатые, с ясным жилкованием, разделение на кориум и перепоночку отсутствует [см. (V)]. Включает около 400 видов.
2. Trichotelocera Reuter 1910 {nom.typ.: Dipsocoris/f=Cryptostemma/g [f:1859; g:1835]. Мелкие (0,5–4 мм). На передних крыльях разделение на кориум и перепоночку может отсутствовать, но иногда область, соответствующая перепоночке, выделяется отсутствием жилок. Включает около 200 видов.
3. Ploteres Latreille 1804a {synn.circ.: Amphibicorisae Dufour 1833 (= Amphibicorises = Amphibicorisa = Amphibiocorisae) = Hydrodromici Burmeister 1835 = Hydrodromica Fieber 1851 = Amphibicoriomorpha Stichel 1955; nom.typ.: Gerris/fg [f:1815; g:1794]}. Адаптированы к обитанию на поверхностной пленке воды; нижняя сторона тела покрыта густыми короткими прилегающими волосками, не смачивающимися водой. У многих представителей разделение переднего крыла на кориум и перепоночку отсутствует или слабо выражено, но у относящихся сюда Ripicolae Amyot & Serville 1843 {nom.typ.: Hebrus/fg} переднее крыло четко разделено на склеротизованный кориум и мембранозную перепоночку, лишенную жилок. Включает более 1 400 видов.
4. Hydrocorisae Latreille 1802b {synn.circ.: Remitarses Dumeril 1805 = Hydroridia Rafinesque 1815 = Cryptocerata Fieber 1851 = Hydrorhynchota Verhoeff 1893a = Euhemiptera Crampton 1916a = Hydrocoriomorpha Stichel 1955; nom.typ.: Nepa/f=Notonecta/g [f:1802; g:L.1758(gen.196)] (incl. Ochterus, Corixa)}. В отличие от остальных Heteroptera, антенны сильно уменьшены, находятся в углублениях на нижней стороне головы под глазами, антеннальные мышцы редуцированы. Большинство Hydrocorisae адаптировано к подводному образу жизни, но дышат атмосферным воздухом, всплывая на поверхность; Aphelocheirus/fg [f:1851; g:1833] не всплывают и дышат растворенным в воде кислородом. Среди Hydrocorisae лишь Gelastocoris/f=Ochterus/g [f:1897; g:1807] ведут наземный прибрежный образ жизни. Большинство Hydrocorisae хищники, лишь Sandaliorrhyncha фитофаги. Включает около 2000 видов. К Hydrocorisae относится таксон Sandaliorrhyncha Börner 1904a {nom.typ.: Corixa/fg [f:1815; g:1762,]}, характеризующийся необычным строением внешнего хоботка, который полностью утратил расчлененность [см. Heteroptera (3)] и неподвижно слит с головой, в отличие от всех прочих Heteroptera.
5. Riparii Burmeister 1835 {nom.typ.: Leptopus/f=Salda/g [f:1835; g:1803]}. Преимущественно прибрежные клопы, большинство с крупными глазами. У самца боковые части II и III тергитов брюшка образуют зажим, удерживающий костальный край переднего крыла самки во время копуляции; при копуляции самец и самка всегда располагаются бок о бок (у некоторых других Heteroptera положение при копуляции бывает таким же, но зажим отсутствует). Включает лишь около 300 видов.
6. Corticicolae Amyot & Serville 1843 {syn.circ.: Phloeobiotica Reuter 1912; nom.typ.: Aradus/fg [f:1837; g:1803]} – подкорники. Тело сильно уплощено, передний край головы с тремя выступами, между которыми прикреплены антенны. Около 800 видов.
7. Trichophora Tullgren 1918 {nom.hier.: Pentatoma/fg [f:1815; g:1789] (incl. Lygaeus, Pyrrhocoris, Coreus, Berytus, Piesma)} характеризуется тем, что на вентральной стороне каждого из III–VII сегментов брюшка обычно имеются по 2 пары трихоботрий [см. Euarthropoda (5с)]; иногда трихоботрий больше двух пар или, наоборот, они утрачены. Включает больше трети всех видов клопов – около 14 000 видов.
8. Cimex/fg (sine Pentatoma; incl. Reduvius, Velocipeda, Miris, Nabis, Tingis). Характеризуется следующими признаками. (1) В отличие от большинства Heteroptera, в половом аппарате самки первичная непарная сперматека утрачена (Pendergrast 1957). (2) Яйцо почти всегда с крышечкой, очерченной ясным кольцевидным швом; микропиле (отверстия для проведения спермы) и аэропиле (отверстия для проведения воздуха) являются разными отверстиями, почти всегда те и другие расположены кольцом кнаружи от крышечки; эмбрион без яйцевого зуба, выходит из яйца, открывая крышечку (тогда как у других клопов на голове эмбриона имеется медиальный яйцевой зуб, служащий для проламывания покрова яйца. Этот таксон включает половину всех видов клопов – около 19 000 видов.
II. Деление Heteroptera на Hydrocorisae и Geocorisae. Таксон Geocorisae Latreille 1817 {synn.circ.: Terrestria Leach 1815a = Gymnocerata Fieber 1851} объединяет всех клопов кроме Hydrocorisae (см. I). В отличие от большинства Hydrocorisae, все Geocorisae обитают в воздушной среде – на суше или на поверхности воды. По мнению разных авторов, Geocorisae является либо парафилетическим, либо голофилетическим таксоном (Mahner 1993).
III. Деление Heteroptera на Hydrocorisae, Ploteres и Geodromica. Таксон Geodromica Fieber 1851 {nom.hier.: Cimex/fg [f:1781; g:L.1758(gen.198)] (sine Notonecta, Gerris)} объединяет всех клопов кроме Hydrocorisae и Ploteres; в отличие от большинства Hydrocorisae и Ploteres, все представители Geodromica обитают на суше. Большинство авторов не признают Geodromica, считая его неоправданно большим парафилетическим таксоном. Однако высказывалось мнение, что Geodromica является голофилетическим таксоном (Pendergast 1957).
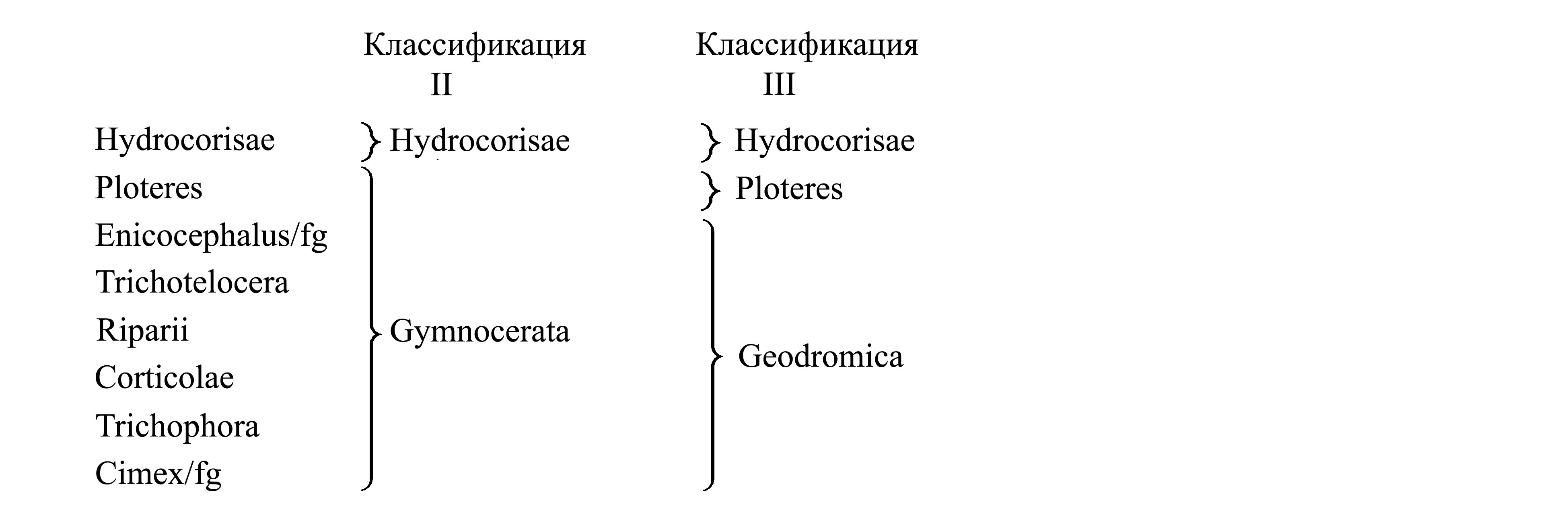
IV. Деление Heteroptera на Archeocorisae и Cimex/fg
1. Плезиоморфон Archeocorisae Carayon 1971 объединяет Enicocephalus/fg, Trichotelocera, Ploteres, Hydrocorisae и Riparii. Характеризуется плезиоморфией: вентральная пахучая железа [см. Heteroptera (7)] открывается на границе заднегруди и брюшка.
2. Cimex/fg (incl. Pentatoma) (= «Geocorisae» sensu Carayon 1971, non Latreille 1817) включает большинство клопов. Характеризуется аутапоморфией: вентральная пахучая железа открывается парой отверстий в основаниях стернальных аподем заднегруди (рис. 9.8.2C).
V. Деление Heteroptera на Panheteroptera и другие группы. Эта классификация основана на следующем представлении о филогении (Stys 1985, Wheeler & Schuh & Bang 1993, Shuh & Slater 1995, et al.):
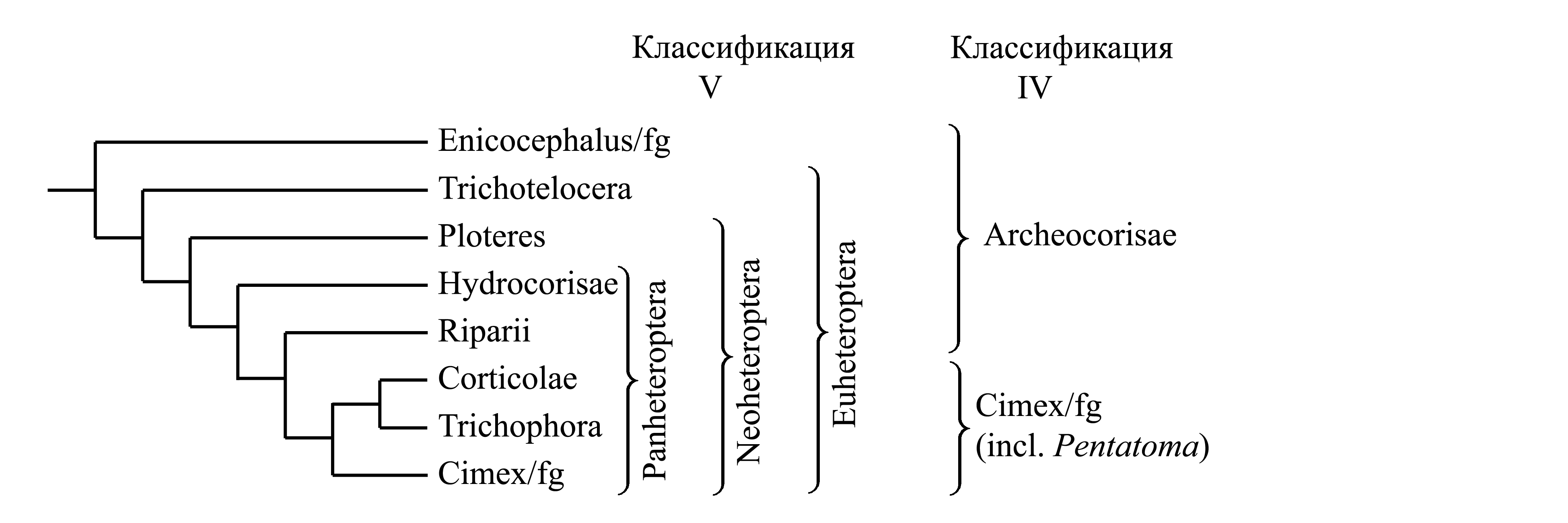
Согласно этой схеме, предложена следующая классификация Heteroptera:
1. Enicocelalus/fg (см. выше).
2. Euheteroptera Stys 1985 {nom.hier.: Cimex/f=Notonecta/g (sine Enicocephalus; incl. Gerris, Ceratocombus)}.
2.1. Trichotelocera (см. выше).
2.2. Neoheteroptera Stys 1985 {nom.hier.: Cimex/f=Notonecta/g (sine Enicocephalus, Ceratocombus; incl. Gerris)}.
2.2.1. Ploteres (см. выше).
2.2.2. Panheteroptera Stys 1985 {nom.hier.: Cimex/f=Notonecta/g (sine Enicocephalus, Ceratocombus, Gerris)} характеризуется тем, что переднее крыло обычно четко разделено на проксимальную утолщенную склеротизованную часть (преклавальная область которой называется кориум) и дистальную тонкую мембранозную часть (перепоночку) так, что при сложенных в покое крыльях перепоночки налегают друг на друга, тогда как кориумы и клавусы не налегают. При этом предполагают, что у тех Panheteroptera, у которых разделение переднего крыла на кориум и перепоночку не выражено, оно утрачено вторично, тогда как у Enicocephalus/fg, Trichotelocera и Ploteres это разделение отсутствует первично (хотя у некоторых Trichotelocera и Ploteres это разделение в той или иной мере намечено).
Эта филогенетическая схема основана на предположении, что разделение переднего крыла на кориум и перепоночку возникло не у общих предков полужесткокрылых [см. Heteropteroidea (3)], а лишь у Panheteroptera. При этом Enicocephalus/fg считается самым примитивным таксоном, сохраняющим перепончатое строение переднего крыла: все крыло состоит из тонкой прозрачной мембраны и сети четких жестких жилок, без признаков уплотнения мембраны и разделения ее на кориум и перепоночку.
Однако строение Enicocephalus/fg сильно модифицировано по сравнению с исходным для Heteropteroidea и для Hemelytrata в целом; в частности, среднегрудь имеет необычную для Hemelytrata форму с сильно увеличенными выпуклыми эпистернами и выпуклым нотумом, что связано с увеличением крыловых мышц непрямого действия [см. Pterygota (4Ab)]. В отличие от других Hemelytrata, самцы Enicocephalus/fg совершают брачный танец в воздухе: они интенсивно летают, постоянно резко меняя направление таким образом, чтобы держаться около одного места; множество самцов, летающих в разных направлениях, образуют единое облако, которое держится над каким-нибудь заметным предметом. Подобный брачный полет, требующий способности к энергичному, долговременному и маневренному полету, встречается у многих комаров (Diptera-Nemocera) и некоторых других хорошо летающих насекомых. Вероятно, перепончатое строение крыльев Enicocephalus/fg является не примитивным, а вторичным и связано с приобретением ими способности к брачному полету. У некоторых Enicocephalus/fg крылья имеются только у молодых имаго и сбрасываются после брачного лета.
* * *