КЛАДОЭНДЕЗИС НАСЕКОМЫХ

![]()
|
КЛАДОЭНДЕЗИС НАСЕКОМЫХ |
|||||||
|
|
|
fg | |||||
Глава IX. СИСТЕМАТИКА PARAMETABOLA
IX-1. Parametabola Crampton 1938
(Somobialia![]() Eucaryota
Eucaryota![]() Metazoa
Metazoa![]() Monostomata
Monostomata![]() Haemataria
Haemataria![]() Peritoneata
Peritoneata![]() Metameria
Metameria![]()
Gnathopoda![]() Euarthropoda
Euarthropoda![]() Mandibulata
Mandibulata![]() Atelocerata
Atelocerata![]() Hexapoda
Hexapoda![]() Amyocerata
Amyocerata![]()
Pterygota![]() Metapterygota
Metapterygota![]() Neoptera
Neoptera![]() Eumetabola
Eumetabola![]() Parametabola)
Parametabola)
(рис. 9.1.1–9.12.4)
|
Названия, однозначно соответствующие по циркумскрипции: — Parametabola Crampton 1938 (non Parametabola Weber 1933). Названия, неоднозначно соответствующие по циркумскрипции (соответствующие также Acercaria): — Panhomoptera Crampton 1918, s.l. — Paraneoptera Martynov 1923, s.l. Иерархическое название: Pediculus/f9=Cicada/g1 [f:1777; g:L.1758(gen.195)] (incl. Zorotypus). |
|
Ранг. Обычно когорта или надотряд.
Название. Наиболее принятым названием этого таксона является Paraneoptera; это соответствует рекомендации Мартынова (1938), который считал, что название Paraneoptera Martynov 1923 предпочтительно, поскольку оно старше названия Parametabola Crampton 1938. Действительно, в статье 1938 г. Мартынов употреблял название Paraneoptera для того же таксона, что и Parametabola, определенно включая сюда Zoraptera. Однако в более ранних публикациях Мартынова (1923 и др.) название Paraneoptera употреблялось в менее определенном значении: Zoraptera не упоминались, и даже положение Thysanoptera было спорным. В настоящее время это дает основание некоторым авторам (Grasse 1949, Grimaldi & Engel 2005 и др.) употреблять название Paraneoptera для таксона, не включающего Zoraptera. На самом деле для таксона, не включающего Zoraptera, имеется старшее и широко признанное название Acercaria Börner 1904a (см. ниже). Для таксона исходно неопределенной циркумскрипции (включающего или не включающего Zoraptera) старшим названием является не Paraneoptera, а Panhomoptera Crampton 1918. Старшим названием, однозначно соответствующим по циркумскрипции рассматриваемому таксону, является Parametabola, поскольку в первой публикации (Crampton 1938) было определенно указано, что Parametabola включает Zoraptera. Употребление названия, однозначно соответствующего по циркумскрипции, в данном случае важно, поскольку в настоящее время существуют принципиальные разногласия о систематическом положении Zoraptera (см. ниже, Zoraptera: «Систематическое положение»). Идентичное название Parametabola было ранее предложено Вебером (Weber 1933), но не для таксона, а для насекомых с определенным типом метаморфоза – для самцов Gallinsecta (но не для самок этих же насекомых).
Аутапоморфии Parametabola
(1) Модификация максилл. Максиллярная лациния [см. Hexapoda (2a)] утратила зубцы и щетинки на медиальной стороне; лишь ее вершина остается заостренной или зазубренной. У Zoraptera лациния изогнута зазубренной вершиной медиально и сохраняет свою исходную функцию в грызущем ротовом аппарате, но погружена в желоб на галеи [см. ниже, Zoraptera (2), рис. 9.1.3C–F]. У Acercaria лациния стала прямой и приобрела способность втягиваться в голову [см. ниже, Acercaria (1)].
(2) Переднемоторность. Крыловой аппарат переднемоторный: в полете переднее крыло ведущее, а заднее сцеплено с ним и двигается пассивно. Крыловая мускулатура [см. Pterygota (4a)] наиболее развита в среднегруди, а в заднегруди она либо развита слабее, либо утрачена. Соответственно, среднегрудь обычно крупнее заднегруди (однако у некоторых Parametabola заднегрудь увеличена в связи с развитием прыгательной мускулатуры задних ног или иной специализации). Переднее крыло обычно (но не всегда) крупнее заднего и часто имеет выраженный торнус – задне-апикальный угол, разделяющий задний край крыла на бази-торнальный и торно-апикальный края; в этом случае передний край заднего крыла примерно равен бази-торнальному краю переднего, так что сцепляясь вместе, оба крыла образуют фигуру цельных очертаний (рис. 9.1.2E, 9.2.1A, 9.7.1A, 9.8.2A, 9.9.1A, 9.10.1A, 9.11.1A).
Сцепка переднего и заднего крыла имеет принципиально разное строение у разных Parametabola: это либо особой формы щетинки по краям обеих пар крыльев [см. ниже, Zoraptera (2) и рис. 9.1.2E]; либо крючковидная щетинка на костальном крае заднего крыла, сцепляющаяся с пучком крепких прямых щетинок на заднем крае переднего крыла [см. ниже, Thysanoptera (6b) и рис. 9.5.1]; либо одна или несколько крючковидных щетинок на костальном крае заднего крыла, сцепляющиеся с подогнутым задним краем переднего крыла [см. ниже, Thysanoptera (6b), Saltipedes (10с), Aphidococca (5) и рис. 9.9.1C]; либо загнутый выступ костального края заднего крыла, сцепляющийся с подогнутым задним краем переднего крыла [см. ниже, Auchenorrhyncha (2) и рис. 9.7.1B]; либо загнутый дорсально костальный край заднего крыла, сцепляющийся с особым замком на вентральной стороне переднего крыла [см. ниже, Panpsocoptera (5) и Heteropteroidea (1), рис. 9.2.1C и 9.8.2B]. Иногда специальный сцепочный аппарат утрачен и опускание задних крыльев обеспечивается тем, что задний край переднего крыла широко налегает на заднее крыло (амплексиформная сцепка) [см. ниже, Scytinelytra (5) и рис. 9.10.1]. Вероятно, исходной для Parametabola или, по крайней мере, для Condylognatha, является сцепка за счет крючковидных щетинок заднего крыла, сцепляющиеся с подогнутым краем переднего крыла: она имеется у примитивных Thysanoptera и у большинства Plantisuga; у некоторых из них, помимо сцепочных щетинок, имеются сходные, но менее изогнутые щетинки, расположенные более проксимально (рис. 9.9.1A); подобные изогнутые проксимальные щетинки имеются также у некоторых Auchenorrhyncha, обладающих иной сцепкой (рис. 9.7.3A). Наличие разных сцепочных аппаратов (функционирующих – рис. 9.5.1 или вестигиальных – рис. 9.7.3A) на одном и том же крыле позволяет понять, как в ходк эволюции Parametabola различные сцепочные аппараты могли заменять друг друга.
Наиболее крупным таксоном в составе Parametabola являются полужесткокрылые (Heteroptera); их передние крылья обычно уплотнены, так что внешне многие клопы напоминают жуков; однако несмотря на поверхностное сходство, специализация крылового аппарата у жуков и клопов прямо противоположная: у жуков это далеко зашедшая заднемоторность, а у клопов – далеко зашедшая переднемоторность, при которой передние крылья совмещают функцию надкрылья с функцией ведущего крыла при полете [см. ниже, Hemelytrata (1с) и Heteropteroidea (3)].
Переднемоторность является неуникальной апоморфией. Независимо от Parametabola переднемоторность, сопровождаемая наличием торнуса и зацепки, возникала в нескольких других группах насекомых [см. Pterygota (4F)] – у поденок и в разных группах насекомых с полным превращением (см. Указатель признаков [2.101]).
(3) Areola postica. На преклавусе переднего крыла [см. Neoptera (1b)] обычно имеется «areola postica» – ячейка, ограниченная ветвями Cu1 и задним краем крыла. Жилка Cu1 в своей дистальной части разделяется на две ветви, впадающие в задний край крыла – Cu1a и Cu1b; передняя из этих ветвей (Cu1a) значительно длиннее задней и изогнута дугообразно выпуклостью вперед, ограничивая спереди «areola postica».
Форма «areola postica» бывает довольно разнообразной: иногда Cu1a так сильно выгнута вперед, что на некотором участке сливается с лежащей впереди M; иногда Cu1a изогнута слабо, и тогда «areola postica» не отличается от краевой ячейки между ветвями Cu1 у других насекомых. Среди Parametabola ячейка «areola postica» отчетливо выражена у многих Eupsocida (рис. 9.2.1A), Saltipedes (рис. 9.9.1A), Zoraptera (рис. 9.1.2A, E), Auchenorrhyncha; у некоторых представителей этих же таксонов она имеет иную форму или совсем не выражена. В связи с сильной модификацией жилкования у Thysanoptera, Gynaptera, Scytinelytra, Gallinsecta и некоторых других, «areola postica» не выражена.
Кроме Parametabola, «areola postica» имеется у вымерших Tetrastigmoptera, которых на этом основании относят к Parametabola (см. ниже, Condylognatha: «Систематическое положение»). По одной из интерпретаций жилкования крыла Diptera, у них также предполагается исходное наличие ячейки, по форме и происхождению соответствующей «areola postica» [ср.: Diptera (2b)].
(4) Утрата шпор. Голень без исходных апикально-внутренних шпор [см. Neoptera (4)]. Иногда на вершине голени бывает развита одна или несколько крепких щетинок, внешне похожих на шпоры. На прыгательных задних ногах Auchenorrhyncha и Saltipedes вершины голеней несут поперечные ряды шиповидных щетинок или неподвижных зубцов, функционально заменяющих шпоры (рис. 9.7.1A); иногда все или некоторые из этих зубцов отчленены и таким образом преобразованы во вторичные шпоры; в частности у Subtericornes-Delphax/fg один зубец преобразован в очень крупную подвижную шпору; у Stridulantes вершина задней голени несет много вторичных шпор. Помимо Parametabola, исходные апикально-внутренние шпоры утрачены также в некоторых других таксонах (см. Указатель признаков [2.48]).
(5) Число тарсомеров. Лапка [исходно пятичлениковая – см. Amyocerata (5)] у имаго состоит не более чем из 3 члеников, у личинки почти всегда не более чем из 2 члеников. В наиболее примитивном случае у личинок всех возрастов лапки 2-члениковые, а у имаго 3-члениковые; при превращении личинки последнего возраста в имаго в первом членике личиночной лапки развивается первый членик имагинальной лапки, а во втором членике личиночной лапки – второй и третий членики имагинальной лапки (рис. 9.2.2B). У многих Parametabola при превращении личинки в имаго не происходит разделения второго членика, в этом случае у имаго лапка 2-члениковая, как и у личинки. В некоторых таксонах число члеников лапки сокращено до одного, либо лапка утрачена. Вторичное увеличение числа члеников лапки произошло только на задних ногах личинок Subtericornes, где их 3, как у имаго.
У Zoraptera лапки личинок и имаго 2-члениковые [см. ниже, Zoraptera (2)]. У сеноедов (Parapsocida и Eupsocida) лапки либо 2-члениковые у личинки и 3-члениковые у имаго (рис. 9.2.2), либо 2-члениковые у личинки и имаго. У Parasita лапки личинок и имаго 2-члениковые или 1-члениковые [см. ниже, Parasita (6), Amblycera (6), Ischnocera (5) и Siphunculata (4)]. У трипсов лапки имаго 2-члениковые или одночлениковые, лапки личинок не отчленены [см. ниже, Thysanoptera (1) и (4)]. У большинства Auchenorrhyncha лапки 2-члениковые у личинки и 3-члениковые у имаго; у некоторых певчих цикад (Auchenorrhyncha-Stridulantes) лапки личинок и имаго 2-члениковые; у Auchenorrhyncha-Subtericornes задние лапки личинок 3-члениковые, как у имаго [см. ниже, Subtericornes (5)]. У Coleorrhyncha лапки личинок и имаго 2-члениковые [см. ниже, Coleorrhyncha (3)]. У большинства Heteroptera лапки либо 2-члениковые у личинки и 3-члениковые у имаго, либо 2-члениковые у личинки и имаго; у некоторых видов лапки 1-члениковые или слиты с голенью. У Heteroptera-Polyctenes/fg1 лапки 2-члениковые у личинки и 3-члениковые у имаго, но голени и лапки имеют вторичные перетяжки, так что лапки средних и задних ног кажутся 4-члениковыми у имаго и 3-члениковыми у личинки. У всех Plantisuga лапки личинки и имаго исходно 2-члениковые [см. ниже, Plantisuga (2)]; у молодых личинок листоблошек и у всех личинок белокрылок лапка слита с голенью [см. ниже, Saltipedes (8) и Scytinelytra (1)]; у некоторых тлей и кокцид лапка утратила расчлененность или отсутствует [см. ниже, Aphidococca (6)].
Помимо Parametabola, несколько сходное увеличение числа члеников лапки до трех происходит в онтогенезе лишь у Microcoryphia и современных стрекоз (Neodonata); у других насекомых лапка либо имеет три членика во всех возрастах (у Embioptera, Plecoptera, Neodermaptera и некоторых других), либо число члеников лапки увеличивается иным образом (у Ephemeroptera, Metabola), либо число члеников более трех (см. Указатель признаков [2.50]: табл. E).
(6) Слияние ганглиев. Все абдоминальные нервные ганглии [см. Hexapoda (1)] слились в один синганглий, находящийся в основании брюшка или в груди; в некоторых группах происходит дальнейшая интеграция нервной системы [см. ниже, Arthroidignatha (2)].
Неуникальная апоморфия, подобное преобразование нервной системы, помимо Parametabola, независимо произошло также в некоторых других группах насекомых.
(7) Уменьшение числа мальпигиевых сосудов. Число мальпигиевых сосудов [стабильное – см. Eumetabola (5)] уменьшено до 6 (т.е. 3 пар). Шесть сосудов имеются только у Zoraptera (Gurney 1938), тогда как у Acercaria число сосудов 4 или меньше [см. ниже, Acercaria (6)]. Те же числа мальпигиевых сосудов встречаются в некоторых других таксонах (см. Указатель признаков [4.7]: табл. F).
(8) Хромосомы голокинетические [см. Eucaryota (2c)]. Голокинетические хромосомы обнаружены как у представителей разных таксонов в составе Acercaria, так и у Zoraptera (Kuznetsova & al. 2002). Однако у некоторых видов выявлены компактные кинетохоры, свойственные монокинетическим хромосомам; в некоторых случаях одна и та же хромосома при разных клеточных делениях (митозе и мейозе) имеет разную длину кинетохора, проявляя свойства то монокинетической, то голокинетической (Wolf 1996). То же у некоторых других насекомых (см. Указатель признаков [4.12.1]).
(9) Уменьшение размеров. Исходным для Parametabola является малый размер тела: среди входящих сюда Zoraptera, Panpsocoptera, Thysanoptera и Plantisuga большинство представителей имеют длину тела 0,5–5 мм, и лишь немногие достигают 1 см и более. Большинство Hemelytrata так же мелкие, но некоторые Subtericornes, Stridulantes и Heteroptera имеют крупные размеры (некоторые особо крупные представители достигают 10 см). Для других групп крылатых насекомых (Ephemeroptera, Odonata, Idioprothoraca, Rhipineoptera и Metabola) характерны преимущественно более крупные размеры, хотя мелкие представители имеются почти в каждой из них (кроме Odonata).
Геологический возраст. Пермь – ныне (Rasnitsyn & Quicke 2002).
Статус Parametabola. Некоторые авторы не признают таксон Parametabola, поскольку считают, что Zoraptera не имеют родства с Acercaria (см. ниже, Zoraptera: «Систематическое положение»).
Классификации Parametabola. Parametabola делятся на голофилетические таксоны Zoraptera и Acercaria. Помимо этого, предлагалось делить Parametabola на парафилетический таксон Lipopteradelphia Crampton 1924 (synn.circ: Panlipoptera Crampton 1924 = Oligonephridia Lameere 1936 = Psocopteroidea Martynov 1938) и голофилетический таксон Arthroidignatha:
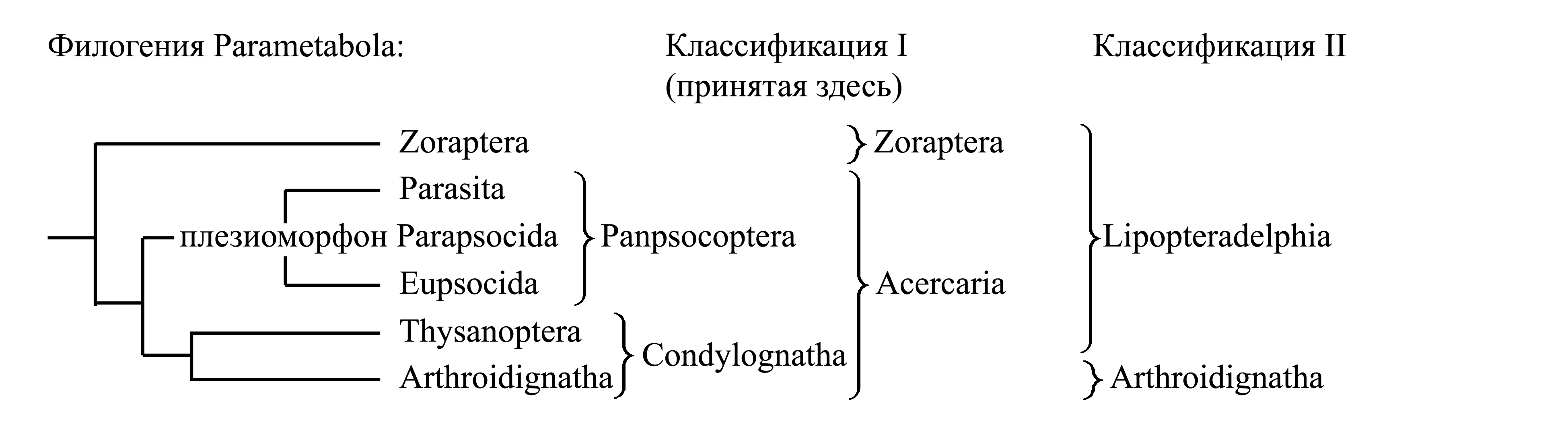
Здесь мы принимаем деление Parametabola на два подчиненных таксона – Zoraptera и Acercaria (стр. 000).
Parametabola, или Pediculus/f9=Cicada/g1
![]() 1. Zoraptera, или Zorotypus/fg1
1. Zoraptera, или Zorotypus/fg1
![]() 2. Acercaria, или Pediculus/f10=Cicada/g2
2. Acercaria, или Pediculus/f10=Cicada/g2
![]() 2.1. Panpsocoptera, или
Pediculus/f11=g1
2.1. Panpsocoptera, или
Pediculus/f11=g1
![]() 2.1.1. Плезиоморфон Parapsocida, или
Atropos/f1=Trogium/g1
2.1.1. Плезиоморфон Parapsocida, или
Atropos/f1=Trogium/g1
![]() 2.1.2. Eupsocida, или Psocus/fg1
2.1.2. Eupsocida, или Psocus/fg1
![]() 2.1.3. Parasita, или Pediculus/f12=g2
2.1.3. Parasita, или Pediculus/f12=g2
![]() 2.1.3.1. Mallophaga, или
Philopterus/f1=Ricinus/g1
2.1.3.1. Mallophaga, или
Philopterus/f1=Ricinus/g1
![]() 2.1.3.1.1. Amblycera, или
Ricinus/f1=g2
2.1.3.1.1. Amblycera, или
Ricinus/f1=g2
![]() 2.1.3.1.2. Ischnocera, или
Philopterus/f2=g1
2.1.3.1.2. Ischnocera, или
Philopterus/f2=g1
![]() 2.1.3.2. Siphunculata, или
Pediculus/f13=g3
2.1.3.2. Siphunculata, или
Pediculus/f13=g3
![]() 2.1.3.3. Rhyncophthirina, или
Haematomyzus/fg(1)
2.1.3.3. Rhyncophthirina, или
Haematomyzus/fg(1)
![]() 2.2. Condylognatha, или Cimex/f3=Cicada/g3
2.2. Condylognatha, или Cimex/f3=Cicada/g3
![]() 2.2.1. Thysanoptera, или
Thrips/fg1–2
2.2.1. Thysanoptera, или
Thrips/fg1–2
![]() 2.2.2. Arthroidignatha, или
Cimex/f4=Cicada/g4
2.2.2. Arthroidignatha, или
Cimex/f4=Cicada/g4
![]() 2.2.2.1. Hemelytrata, или
Cimex/f5=Cicada/g5
2.2.2.1. Hemelytrata, или
Cimex/f5=Cicada/g5
![]() 2.2.2.1.1. Auchenorrhyncha,
или Cicada/f1=g6
2.2.2.1.1. Auchenorrhyncha,
или Cicada/f1=g6
![]() 2.2.2.1.1.1. Subtericornes,
или Fulgora/fg1
2.2.2.1.1.1. Subtericornes,
или Fulgora/fg1
![]() 2.2.2.1.1.2. Euhomoptera, или
Cicada/f2=g7
2.2.2.1.1.2. Euhomoptera, или
Cicada/f2=g7
![]() 2.2.2.1.2. Heteropteroidea,
или Cimex/f6=Notonecta/g1
2.2.2.1.2. Heteropteroidea,
или Cimex/f6=Notonecta/g1
![]() 2.2.2.1.2.1. Coleorrhyncha,
или Peloridium/fg1
2.2.2.1.2.1. Coleorrhyncha,
или Peloridium/fg1
![]() 2.2.2.1.2.2. Heteroptera, или
Cimex/f7=Notonecta/g2
2.2.2.1.2.2. Heteroptera, или
Cimex/f7=Notonecta/g2
![]() 2.2.2.1.2.3. †Progonocimex/fg1
2.2.2.1.2.3. †Progonocimex/fg1
![]() 2.2.2.2. Plantisuga, или
Aphis/fg1
2.2.2.2. Plantisuga, или
Aphis/fg1
![]() 2.2.2.2.1. Psyllaleyroda, или
Psylla/fg1
2.2.2.2.1. Psyllaleyroda, или
Psylla/fg1
![]() 2.2.2.2.1.1. Saltipedes, или
Psylla/fg2
2.2.2.2.1.1. Saltipedes, или
Psylla/fg2
![]() 2.2.2.2.1.2. Scytinelytra,
или Aleyrodes/fg1
2.2.2.2.1.2. Scytinelytra,
или Aleyrodes/fg1
![]() 2.2.2.2.2. Aphidococca,
или Aphis/fg2
2.2.2.2.2. Aphidococca,
или Aphis/fg2
![]() 2.2.2.2.2.1. Gynaptera, или
Aphis/fg3
2.2.2.2.2.1. Gynaptera, или
Aphis/fg3
![]() 2.2.2.2.2.2. Gallinsecta, или
Coccus/fg1
2.2.2.2.2.2. Gallinsecta, или
Coccus/fg1