КЛАДОЭНДЕЗИС НАСЕКОМЫХ

![]()
|
КЛАДОЭНДЕЗИС НАСЕКОМЫХ |
|||||||
|
|
|
fg | |||||
IX-1.2.2.2.1. Hemelytrata Fallén 1829
(Somobialia![]() Eucaryota
Eucaryota![]() Metazoa
Metazoa![]() Monostomata
Monostomata![]() Haemataria
Haemataria![]() Peritoneata
Peritoneata![]() Metameria
Metameria![]()
Gnathopoda![]() Euarthropoda
Euarthropoda![]() Mandibulata
Mandibulata![]() Atelocerata
Atelocerata![]() Hexapoda
Hexapoda![]() Amyocerata
Amyocerata![]()
Pterygota![]() Metapterygota
Metapterygota![]() Neoptera
Neoptera![]() Eumetabola
Eumetabola![]() Parametabola
Parametabola![]() Acercaria
Acercaria![]() Condylognatha
Condylognatha![]()
Arthroidignatha![]() Hemelytrata)
Hemelytrata)
(рис. 9.7.1–9.8.2)
(рис. 9.7.1,
9.7.2,
9.7.3, 9.8.1,
9.8.2)
|
Названия, однозначно соответствующие по циркумскрипции: — Hemelytrata Fallén 1829; — Euhemiptera Zrzavy 1990b (non Euhemiptera Crampton 1916a); — Tymbalia Wessel & al. 2014. Иерархическое название: Cimex/f3=Cicada/g5 [f:1781(ord.3); g:L.1758(gen.195)] (sine Aphis). |
|
Ранг. Ниже или выше отряда.
Аутапоморфии Hemelytrata
(1) Складывание крыльев. Крылья, будучи в покое сложены на спине [см. Neoptera (1)], плотно прижаты к телу [см. (1d)] благодаря особенностям строения груди [см. (1a)], брюшка [см. (1b)] и передних крыльев [см. (1c)]. У некоторых Hemelytrata способность плотно складывать крылья вторично утрачена [см. (1e)].
Часто передние крылья склеротизованы и, будучи плотно сложены на спине, выполняют функцию надкрыльев; при этом они могут сохранять ясно выраженное жилкование, но в случаях, когда мембрана сильно склеротизована и утолщена (у некоторых Heteroptera) жилки бывают плохо выражены или совсем не видны. Несмотря на это, передние крылья всегда остаются ведущими в полете [см. Parametabola (2)]. У некоторых Hemelytrata крылья укорочены или утрачены [см. Neoptera (1e)].
(1 a) Особенности строения тергитов груди. Тергит среднегруди (мезонотум) имеет особый щиток (scutellum) – четко очерченный задний участок в форме треугольника, обращенного вершиной назад; поверхность щитка приподнята над лежащей позади и латеральнее его поверхностью груди и брюшка; задне-латеральные края щитка имеют желобки, обращенные назад-латерально (рис. 9.7.3A) [следует упомянуть, что термин «щиток» (scutellum) употребляется в двух разных значениях: исходно он применялся только для данного специализированного участка среднегруди Hemelytrata, а позже (Audouin 1824) стал использоваться также для большего по размеру участка среднегруди и заднегруди любых крылатых насекомых – см. Pterygota (4a)].
Тергит переднегруди (передеспинка, или пронотум) обычно простирается назад в виде большой лопасти, которая закрывает значительную часть мезонотума, оставляя открытым только щиток. При такой форме пронотума кажется, что насекомое имеет длинную переднегрудь и короткую среднегрудь (рис. 9.7.3A), напоминая этим жуков; в действительности, наоборот: переднегрудь короткая, а среднегрудь увеличена и вмещает мощную крыловую мускулатуру (рис. 9.7.3D), как это свойственно переднемоторным насекомым [см. Parametabola (2)]. У некоторых Hemelytrata (Stridulantes, многие Subtericornes и др.) лопасть пронотума и/или щиток мезонотума не развиты [см. (1e)].
(1 b) Особенности строения брюшка. III–VIII уромеры имеют резко выраженный латеральный край, который выступает в виде гребня и отделяет дорсальную сторону, образованную тергитами, от вентральной стороны, образованной стернитами и плейритами (у Auchenorrhyncha более выпуклой обычно является дорсальная сторона, у Heteropteroidea – обычно вентральная). Дыхальца III–VIII уромеров расположены на вентральной стороне. I и II уромеры имеют меньшие размеры и иное строение; дыхальца I уромера расположены на дорсальной стороне, дыхальца II уромера расположены по-разному в разных таксонах: у Auchenorrhyncha и Coleorrhyncha – на дорсальной стороне (рис. 9.7.1A, 9.8.1E), у Heteroptera – на вентральной стороне (рис. 9.8.2A). Обычно развиты все 8 пар абдоминальных дыхалец [помимо двух пар межсегментарных дыхалец на груди – см. Amyocerata (2)]; лишь у некоторых Heteroptera часть абдоминальных трахеостий замкнута, так что число функционирующих дыхалец может быть сокращено.
Резкий латеральный край брюшка встречается также у многих других насекомых, но он не характерен для крылатых имаго Plantisuga.
(1 c) Особенности строения переднего крыла. Исходно переднее крыло склеротизовано, так что совмещает в себе функцию ведущего крыла при полете [см. Parametabola (2)] и функцию надкрылья, защищающей заднее крыло в сложенном состоянии [у многих клопов передние крылья склеротизованы наполовину – см. ниже, Heteropteroidea (3)]. Клавальный желоб [отделяющий клавус от преклавуса – см. Neoptera (1b)] представляет собой прямую борозду, которая позволяет крылу перегибаться по ней (рис. 9.7.1, 9.7.3A, 9.8.2). Задний край крыла в области клавуса [см. (2a)] выступает в виде тупого угла, так что проксимальнее этого угла имеется прямой базиклавальный край, а дистальнее – прямой дистиклавальный край. Базиклавальный край продолжается несколько проксимальнее-дорсальнее места причленения к крылу третьего аксиллярного склерита, образуя базиклавальный козырек. У некоторых представителей эти особенности крыльев утрачены [см. (1e)].
(1 d) Складывание крыльев. При складывании переднего крыла его прямой базиклавальный край плотно прижимается к задне-латеральному краю щитка и закрепляется в его желобке [см. (1a)], а прямой дистиклавальный участок сходится по средней линии с соответствующим участком парного ему крыла (рис. 9.7.3A). Костальный край переднего крыла прижимается к латеральному гребню брюшка [см. (1b)] или к дорсальной стороне брюшка медиальнее латерального гребня. Благодаря этому по крайней мере дыхальца III–VIII уромеров оказываются не закрытыми крыльями (в отличие от Eleuterata, у которых, наоборот, абдоминальные дыхальца расположены дорсально и открываются в субэлитральную полость). Задняя часть заднего крыла [вместе с подвернутой под нее инверталярной лопастью – см. (2b)] при складывании крыла заходит за среднюю линию тела, так что задние крылья налегают друг на друга (Fallén 1814).
(1 e) Разнообразие в складывании крыльев. Исходно (у большинства Auchenorrhyncha) передние крылья складываются либо кровлеобразно (тогда их преклавальные области сходятся медиально своими задними краями, являющимися прямыми продолжениями дистиклавальных краев), либо облегающее (тогда дистальные части их преклавальных областей несколько налегают друг на друга). В обоих случаях задние крылья складываются облегающе, налегая друг на друга своими задними частями с подвернутыми инверталярными лопастями. У большинства Heteropteroidea крылья складываются плоско; при этом задние крылья налегают так же как и у Auchenorrhyncha, а передние крылья широко налегают друг на друга дистальными частями (рис. 9.8.1A).
У некоторых цикадовых, фульгороидов и полужесткокрылых те или иные из названных выше особенностей строения груди и/или крыльев отсутствуют: щиток может не иметь желобков, переднеспинка может быть без задней лопасти, клавус переднего крыла может быть без выступающего угла, клавальные козырьки могут не выступать, задние крылья при складывании могут не налегать друг на друга; в некоторых случаях крылья вообще утратили способность складываться в покое. Может сложиться ошибочное мнение, что неспособность плотно складывать крылья является примитивной чертой. В частности, у певчих цикад (Stridulantes) передние крылья в покое не способны плотно прижиматься к щитку и друг к другу, среднеспинка не образует четко очерченного щитка, а задние крылья не налегают друг на друга; однако явное родство певчих цикад с Tettigarcta/fg (см. ниже, Euhomoptera: «Классификация») и наличие у Tettigarcta/fg такого же способа складывания крыльев, как у многих других Auchenorrhyncha, говорит о том, что способность плотно складывать крылья является исходной.
(2) Модификация крыльев. На крыльях обеих пар жилки и складки взаиморасположены таким образом, что клавус (область между клавальным желобом и инверталярной складкой) имеет 2 жилки [см. (2a)], а инверталяре (переворачивающаяся область позади инверталярной складки) на переднем крыле не имеет жилок, а на заднем крыле увеличена и имеет одну жилку [см. (2b)].
(2 a) Клавус. Клавус [см. Neoptera (1b)] на крыльях обеих пар исходно имеет 2 продольные клавальные жилки, обычно обозначаемые как Pcu и V1 (или, соответственно, 1A и 2A) (тогда как у других Parametabola между клавальным желобом и задним краем крыла имеется не более одной жилки). На переднем крыле обе эти жилки идут на расстоянии от клавального желоба и от сцепочного подгиба заднего края [см. (3)]; у Subtericornes, Coleorrhyncha и некоторых Heteroptera эти жилки дистально сливаются, образуя Y-образную фигуру; у Euhomoptera эти две жилки впадают в задний край крыла порознь (рис. 9.7.3A) [см. ниже, Euhomoptera (4)]. На заднем крыле обе клавальные жилки впадают в край крыла порознь [позади них имеется еще одна жилка на подгибающейся части крыла – см. (2b)].
У различных представителей Hemelytrata те или иные жилки исчезают; в частности, на переднем крыле жилки иногда исчезают в связи с сильной склеротизацией мембраны крыла [см. (1)], а на заднем крыле – в связи с десклеротизацией жилок.
(2 b) Инверталяре заднего крыла. Заднее крыло [способное сцепляться с передним – см. (3) и Parametabola (2)] имеет расширенную инверталярную лопасть, переворачивающуюся при складывании крыла [см. Neoptera (1a)]; при этом инверталярная складка проходит сразу позади V1 (или 2A), а на инверталярной лопасти имеется единственная жилка V2 (или 3A) (рис. 9.7.1A, 9.8.2A).
Высказывалось мнение, что наличие инверталярной лопасти у Hemelytrata является плезиоморфией, поскольку еще более крупная инверталярная лопасть (или анальный веер) имеется у многих примитивных новокрылых [ср.: Rhipineoptera (1)]. Однако, вероятнее всего, инверталярная лопасть у Hemelytrata представляет собой новообразование: у Zoraptera, Parapsocida, Eupsocida и Plantisuga ее нет, так что, видимо ее не было и у общего предка Parametabola. В отличие от Rhipineoptera, у Hemelytrata инверталярная лопасть, независимо от ее размеров, имеет лишь одну жилку и не складывается веерообразно.
(3) Крыловая сцепка. Сцепка заднего крыла с передним [см. Parametabola (2)] обеспечивается дорсальным загибом переднего края заднего крыла; сцепочные щетинки в этом месте отсутствуют (в отличие от Plantisuga и Thysanoptera – см. Указатель признаков [2.152]). Крючковидные щетинки на заднем крыле либо совсем отсутствуют, либо имеются лишь в проксимальной части переднего края и не обеспечивают сцепки. Конструкция сцепки сильно различается у разных Hemelytrata: у цикадовых короткий загиб (крюк) на заднем крыле цепляется за длинный загиб на переднем крыле [см. ниже, Auchenorrhyncha (2) и рис. 9.7.1B]; у клопов, наоборот, длинный загиб на заднем крыле цепляется за короткий замок на переднем [см. ниже, Heteropteroidea (1) и рис. 9.8.2B].
(4) Особенности антенн. Флагеллум антенны утратил способность к увеличению числа члеников путем деления проксимальных члеников [см. Amyocerata (1)]. В процессе онтогенеза флагеллум либо сохраняет постоянное число члеников, либо меняет свое расчленение, но не путем деления проксимальных члеников [см. ниже, Auchenorrhyncha (1), Coleorrhyncha (2) и Heteroptera (4)].
(5) Орган Эванса. На голове имеется парный орган Эванса, представляющий собой плакоидную сенсиллу [производную щетинки – см. Euarthropoda (5)], которая погружена в камеру, открывающуюся наружу узким отверстием [«a finger-like lobe contained in a pit» (Evans 1973)]. У разных представителей Auchenorrhyncha и Coleorrhyncha пара органов Эванса расположена либо на максиллярных пластинках [см. Arthroidignatha (1b)], либо дорсальнее их и вентральнее антенн. У Heteroptera органов Эванса нет (Bourgoin 1986).
(6) Аролий. На претарсусе между двумя коготками имеется пузыревидный аролий [см. Pterygota (5)]. Такой аролий имеется у личинок и имаго Coleorrhyncha и большинства Auchenorrhyncha; среди Auchenorrhyncha лишь у личинок цикад с их специализированными коготками аролий утрачен, а у имаго вестигиальный (см. ниже, Euhomoptera: «Классификация»: Cicada/f3=g8). У клопов аролий утрачен [см. ниже, Heteroptera (7)].
(7) Гениталии самца. У самца парамеры [см. Condylognatha (2)] снабжены аподемами, вдающимися вглубь IX уромера, благодаря чему способны выдвигаться и погружаться (рис. 9.4.2A–C, 9.8.2A).
В отличие от Saltipedes и Scytinelytra, у Hemelytrata парамеры не сочленены непосредственно с задним краем IX стернита брюшка (хотя бывают приближены к нему). Вероятно, исходным для Hemelytrata является строение гениталий, общее для большинства Auchenorrhyncha-Euhomoptera и Heteropteroidea-Coleorrhyncha: парамеры подвижно сочленены с основанием пениса и, помимо мышц, идущих к IX стерниту брюшка, сохраняют мышцы, идущие к основанию пениса; пенис не имеет непосредственных сочленений со склеритами IX или X сегментов брюшка, так что способен в той или иной мере втягиваться. У Heteroptera парамеры утратили сочленение с пенисом и мышцы, идущие к пенису [см. ниже, Heteroptera (10)]. У Euhomoptera-Stridulantes и некоторых Heteroptera парамеры утрачены.
Пенис бывает цельным (у Stridulantes), либо подразделен на подвижно сочлененные фаллобазу и эдеагус (у Cercopis/fg и Heteroptera), либо имеет отчлененный склерит фаллобазы, сочлененный с парамерами (у Membracis/fg); у фульгороидов гениталии усложнены особым образом [см. ниже, Subtericornes (7)]. Семяизвергательный канал непарный и обычно открывается одним гонопором [см. Hexapoda (12)], но у отдельных представителей неродственных друг другу таксонов Euhomoptera пенис и проходящий в нем семяизвергательный канал раздвоен на вершине так, что имеется пара половых отверстий (Емельянов 1987). Тергит и стернит IX уромера обычно слиты в цельное кольцо, или генитальную капсулу, называемую пигофор (pygophorus), или пигофер (pygofer); лишь у некоторых Euhomoptera-Cercopis/fg1 и Euhomoptera-Membracis/fg1 тергит и стернит IX уромера вторично разделены (Емельянов 1987).
(8) Анальная трубка. Последний (десятый) сегмент брюшка [см. Hexapoda (8)] преобразован в анальную трубку: он утончен и разделен на два вторичных членика, так что анальное отверстие находится на вершине второго членика. Каждый из двух члеников анальной трубки имеет склеротизованную цилиндрическую поверхность, а членики подвижно соединены эластичной мембраной, так что дистальный членик способен частично или полностью втягиваться в проксимальный (рис. 9.4.2A –C). У Auchenorrhyncha оба членика хорошо выражены, а к вершине дистального членика вентральнее анального отверстия подвижно причленен анальный столбик – непарный отросток с желобом на дорсальной стороне (рис. 9.7.1A). У Coleorrhyncha дистальный членик имеет вид тонкого склеротизованного цилиндра и способен полностью втягиваться в проксимальный членик (рис. 9.8.1C). У Heteroptera дистальный членик маленький, мембранозный (рис. 9.8.2A), в покое втянут в проксимальный и незаметен. Анальная трубка способствует тому, что выделяемая из анального отверстия медвяная роса [см. Arthroidignatha (1f)] удаляется, не загрязняя тело.
В отличие от Hemelytrata, у Plantisuga десятый уромер не только не расчленен, но, наоборот, полностью слит с тергитом девятого уромера.
Некоторые авторы (Snodgrass 1935 и др.) считают проксимальный членик анальной трубки десятым сегментом брюшка, а дистальный членик – исходным одиннадцатым сегментом брюшка; при этом считают, что расчлененная анальная трубка Hemelytrata является плезиоморфией, а отсутствие такого расчленения у других насекомых – апоморфией. Эта интерпретация связана с ошибочным представлением об исходном наличии у насекомых XI сегмента брюшка [см. Hexapoda (8b)].
Плезиоморфии и изменчивые признаки Hemelytrata
(9) Яйцеклад. В отличие от других Parametabola, у самки обычно сохраняется яйцеклад, состоящий из 3 пар створок, сочлененных парой гонангулов [см. Amyocerata (7)]; при этом пара VIII кинетапофизов и слитые между собой IX кинетапофизы сцеплены олистетерами и образуют режущий яйцеклад, а придатки IX уромера, считающиеся стилигерами и/или стерностилями, образуют ножны характерного строения [см. Eumetabola (1)] (рис. 9.7.1A). У некоторых Heteroptera (Hydrocorisae, Trichophora и др.) ножны яйцеклада утрачены; у некоторых Subtericornes и некоторых Heteroptera яйцеклад вестигиальный или утрачен.
Геологический возраст. Пермь – ныне (Rasnitsyn & Quicke 2002).
Статус Hemelytrata. В некоторых классификациях таксон Hemelytrata отсутствует (см. Arthroidignatha: «Классификации» I).
Классификации Hemelytrata
I. Деление Hemelytrata на Auchenorrhyncha и Heteropteroidea. Об апоморфиях этих таксоном см. ниже.
II. Деление Hemelytrata на Euhomoptera и Neohemiptera. Таксон Euhomoptera рассматривается нами как подчиненный в составе Auchenorrhyncha – см. ниже. Таксон Neohemiptera Sorensen et al. 1995 объединяет Subtericornes и Heteropteroidea; по мнению некоторых авторов он является голофилетическим и характеризуется следующими признаками.
(1) Мандибулярные пластинки [см. Arthroidignatha (1a)] в своей дорсальной части не отделены от клипеуса и лба [см. ниже, Subtericornes (1)].
(2) На переднем крыле жилки клавуса Pcu и V1 слиты апикально, образуя Y-образную фигуру [см. Hemelytrata (2a) и ниже, Subtericornes (9)].
(3) Периферическая мембрана на заднем крыле отсутствует [см. Auchenorrhyncha (7)].
(4) Фильтрационная камера [см. Arthroidignatha (3)] отсутствует (то же у части Gynaptera).
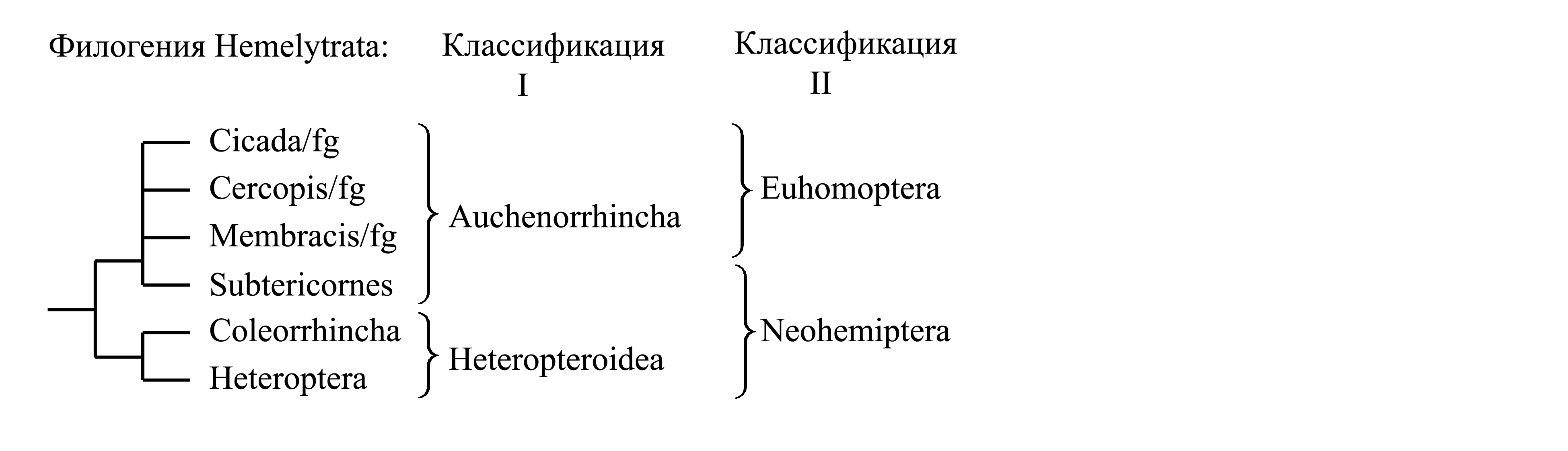
Здесь мы принимаем деление Hemelytrata на два подчиненных таксона – Auchenorrhyncha и Heteropteroidea (стр. 000).
Hemelytrata, или Cimex/f3=Cicada/g5
![]() 1.
Auchenorrhyncha,
или Cicada/f1=g6
1.
Auchenorrhyncha,
или Cicada/f1=g6
![]() 1.1. Subtericornes, или
Fulgora/fg1
1.1. Subtericornes, или
Fulgora/fg1
![]() 1.2. Euhomoptera, или Cicada/f2=g7
1.2. Euhomoptera, или Cicada/f2=g7
![]() 2. Heteropteroidea, или
Cimex/f4=Notonecta/g1
2. Heteropteroidea, или
Cimex/f4=Notonecta/g1
![]() 2.1. Coleorrhyncha, или
Peloridium/fg1
2.1. Coleorrhyncha, или
Peloridium/fg1
![]() 2.2. Heteroptera, или Cimex/f5=Notonecta/g2
2.2. Heteroptera, или Cimex/f5=Notonecta/g2
![]() 3. †Progonocimex/fg1
3. †Progonocimex/fg1