КЛАДОЭНДЕЗИС НАСЕКОМЫХ

![]()
|
КЛАДОЭНДЕЗИС НАСЕКОМЫХ |
|||||||
|
|
|
fg | |||||
X-1.3.2. Enteracantha Kluge 2010 – скорпионницы и блохи
(Somobialia![]() Eucaryota
Eucaryota![]() Metazoa
Metazoa![]() Monostomata
Monostomata![]() Haemataria
Haemataria![]() Peritoneata
Peritoneata![]() Metameria
Metameria![]()
Gnathopoda![]() Euarthropoda
Euarthropoda![]() Mandibulata
Mandibulata![]() Atelocerata
Atelocerata![]() Hexapoda
Hexapoda![]() Amyocerata
Amyocerata![]()
Pterygota![]() Metapterygota
Metapterygota![]() Neoptera
Neoptera![]() Eumetabola
Eumetabola![]() Metabola
Metabola![]() Mecopteriformia
Mecopteriformia![]() Enteracantha)
Enteracantha)
(рис. 10.8.1–10.13.3)
(рис. 10.8.1,
10.8.2,
10.9.1,
10.9.2, 10.10.1,
10.10.2,
10.11.1,
10.11.2, 10.12.1,
10.12.2,
10.12.3,
10.13.1,
10.13.2,
10.13.3)
|
Название, однозначно соответствующее по циркумскрипции: — Enteracantha Kluge 2010a. По циркумскрипции также соответствует: — «Mecopterida» sensu Grimaldi & Engel 2005 (non Pearce 1936, nec Boudreaux 1979b, nec Whiting et al. 1997) Иерархическое название: Panorpa/fg1 [f:1802; g:L.1758(gen.221)] (incl. Pulex, Boreus, Bittacus, Nannochorista). |
|
Ранг. Отряд или выше.
Аутапоморфии Enteracantha
(1) Аканты. У имаго внутренняя стенка провентрикулюса [см. Gnathopoda (3d)] несет многочисленные (порядка нескольких сотен) кутикулярные выросты – аканты; каждая эпителиальная клетка провентрикулюса образует одну аканту [так что аканты принципиально отличаются от щетинок – см. Euarthropoda (5d)]; аканта бывает длиной 50–150 μm, так что бóльшая часть полости провентрикулюса заполнена плотно сомкнутыми акантами (рис. 10.8.1) (Richards & Richards 1969).
Функционального значения аканты, по-видимому, не имеют, их наличие не коррелирует с составом потребляемой пищи. Высказывалось предположение, что у блох аканты служат для разрушения кровяных клеток, однако это не так, и эритроциты проходят в среднюю кишку неповрежденными (Ващенок 1988); у блох Tunga penetrans (Linnaeus 1758), паразитирующих в толще кожи хозяина, аканты имеются только у молодых самок, но исчезают после начала питания. Наличие акант в провентрикулюсе – очень консервативный признак: они имеются у всех скорпионниц и блох. Так что это признак позволяет не только выявить принадлежность всех скорпионниц и блох к одной филогенетической ветви, но и сделать вывод, что к этой ветви не относятся насекомые, не имеющие акант. В частности, сюда не относятся Diptera, которых некоторые авторы считают потомками скорпионниц и/или предками блох.
(2) Рострум. У имаго голова исходно вытянута, образуя рострум (rostrum): клипеус [см. Amyocerata (3)] и латеральные участки головной капсулы (к которым причленены мандибулы) вытянуты, образуя переднюю часть рострума, а лабио-максиллярный комплекс [см. Mecopteriformia (2)] вытянут, образуя заднюю часть рострума (рис. 10.9.1A); передние тенториальные ямки находятся позади вытянутого клипеуса, так что весь рострум вытянут впереди тенториума (рис. 10.8.1A–B). Верхняя губа неподвижно слита с клипеусом, фронто-лабральные мышцы утрачены; в некоторых таксонах, у которых рострум утрачен (Nannomecoptera, Raphioptera-Caurinus/fg, Aphaniptera и др.), верхняя губа может быть отделена от клипеуса швом, но и в этих случаях она не имеет мышц. Мандибулы причленяются на вершине рострума по бокам от неподвижной верхней губы. Свободные части максилл (лацинии, галеи и 5-члениковые щупики) и свободная часть нижней губы [прементум со щупиками – см. (5)] также отходят от вершинной части рострума. У представителей с длинным рострумом максилла может иметь аподему, которая начинается от основания лацинии, тянется в проксимальном направлении вдоль стипеса (входящего в состав вентральной стенки рострума) и служит местом прикрепления кранио-лациниальной мышцы [см. Hexapoda (2)]; мышцы, идущие из кардо и стипеса максилл [исходно прикрепленные к передним рукам тенториума – см. Amyocerata (3)] могут быть частично или полностью перемещены на ту часть головной капсулы, которая входит в состав рострума (Hepburn 1969); у блох (не имеющих ни рострума, ни тенториума) они крепятся к головной капсуле [см. ниже, Aphaniptera (1a) и (1b)].
Длинный хорошо выраженный рострум имеется у многих Scorpiomuscae (Panorpa/fg3, Notiothauma/fg, Apteropanorpa/fg), у Metamecoptera и у большинства Raphioptera (Boreus/fg2). У некоторых Enteracantha рострум подвергся редукции: у части Scorpiomuscae (Merope/fg1, Panorpodes/fg), у Nannomecoptera, у Raphioptera-Caurinus/fg и у блох (Aphaniptera) рострум укорочен или вообще не выражен (рис. 10.11.1, 10.13.1A).
Несмотря на модификацию головы, ротовой аппарат может сохранять исходную грызущую функцию: у Scorpiomuscae, Metamecoptera и Raphioptera имеются вполне развитые грызущие мандибулы [со сближенными передним и задним мыщелками – см. Metapterygota (2)]; у наннохорист и блох ротовой аппарат модифицирован [см. ниже, Nannomecoptera (2) и Aphaniptera (1b)].
Данные особенности присущи только имаго, тогда как у личинки голова всегда без рострума, верхняя губа отчленена (рис. 10.9.2B, 10.12.3).
(3) Модификация пронотума. У имаго тергит переднегруди имеет характерную форму: его задне-латеральные углы вытянуты назад так, что сочленяются с передними краями эпистернов среднегруди. Стеноторакальные дыхальца [исходно находящиеся на мембране между переднегрудью и среднегрудью – см. Amyocerata (2)] бывают расположены на этих склеротизованных углах пронотума (рис. 10.12.1A) или на мембране дорсальнее их (рис. 10.9.1A, 10.11.1). У блох вытянутые углы пронотума, сочлененные с эпистенрами среднегруди и несущие стеноторакальные дыхальца, преобразованы в обособленные склериты, подвижно причленные к месту соединения пронотума и плейрита (рис. 10.13.1A, 10.13.2B) [см. ниже, Aphaniptera (1a)].
(4) Утрата дистальной части личиночной ноги и исходное строение личинок. У личинки ноги вестигиальные: каждая нога утратила всю дистальную часть, включая коленное сочленение, голень, лапку [исходно одночлениковую – см. Metabola (1Cc)] и коготок [исходно единственный – см. Mecopteriformia (4)]; так что нога состоит только из тазика, вертлуга и бедра, являющегося последним члеником ноги (рис. 10.9.2C, 10.11.2A–D). У личинок Raphioptera ноги имеют еще более упрощенное строение (рис. 10.12.3), а у личинок Aphaniptera ноги совсем утрачены.
Некоторые авторы ошибочно считали, что у относящихся сюда Nannomecoptera личиночная нога имеет 2-члениковую лапку и два коготка; в этом случае тазик принимали за бедро, вертлуг за голень, бедро за лапку, а имеющиеся на бедре кутикулярные крючки – за коготки (Pilgrim 1972). Расположение мышц позволяют правильно гомологизировать членики ноги (рис. 10.11.2A –D) (Kluge 2004): те членики, к основанию которых мышцы подходят прямо из туловища, являются тазиком и вертлугом, так что следующий после вертлуга членик (самый дистальный) является бедром. Крючья, сидящие на вершине ноги у Nannomecoptera, не являются истинными коготками и не относятся к претарсусу, поскольку у них отсутствует унгуитрактор с мышцами-сгибателями, но имеется мышца-разгибатель, которой не бывает у претарсуса насекомых [см. Atelocerata (3)].
Вероятно, исходно личинки Enteracantha приспособлены для ползанья в почве и подстилке: такая личинка имеет эластичное удлиненное тело, способное волнообразно накатываться вперед; дистальные отделы ног (соответствующие бедрам) мягкие, способны вытягиваться и сжиматься; на вершине X (последнего) сегмента брюшка имеются выпячивающиеся отростки [пигопод – см. Metabola (1A)]; I–VIII сегменты брюшка несут по паре ложноножек (рис. 10.9.2B и 10.10.2B). Ложноножки имеют простую коническую форму и не имеют мышц; они расположены не в общий ряд с грудными ногами, а значительно медиальнее (так что не гомологичны ложноножкам Hymenoptera и Neolepidoptera, расположенным в один ряд с ногами) (см. Указатель признаков [3.44]). При ползании личинка приподнимает переднюю часть тела, вытягивает ее вперед и опускается на нее, вытянув вперед ноги; затем она сокращает переднюю часть тела и ноги, подтягиваясь вперед. Личинки такого строения свойственны некоторым Scorpiomuscae (Panorpa/fg3, Chorista/fg) и Metamecoptera [см. ниже, Scorpiomuscae (6) и Metamecoptera (3)]. У Scorpiomuscae-Panorpodes/fg1 и Raphioptera личинки менее подвижны, С-образно изогнуты и без ложноножек (рис. 10.12.3) [см. ниже, Raphioptera (4)]. Личинки Nannomecoptera перешли к обитанию в воде, имеют длинное извивающееся тело без ложноножек, но с парой крючков на десятом сегменте брюшка (рис. 10.11.2); их ноги приобрели вторичные когтевидные крючья, но состоят из того же редуцированного набора члеников, что и ноги подстилочных личинок Scorpiomuscae и Metamecoptera (что свидетельствует об их происхождении от подстилочных предков) [см. ниже, Nannomecoptera (1)]. У личинок блох ноги совсем утрачены, ложноножки также отсутствуют [см. ниже, Aphaniptera (7)].
Несмотря на редукцию ног, личинки большинства Enteracantha активно перемещаются в поисках пищи: у всех Enteracantha личинки сохраняют грызущий ротовой аппарат с развитыми мандибулами; личинки Scorpiomuscae, Metamecoptera и Nannomecoptera хищные, личинки Calyptroptera преимущественно растительноядные или детритоядные.
Признаки Enteracantha неясного филогенетического значения
(5) Утрата глосс и параглосс. У имаго и личинки нижняя губа [не имеющая парных глосс – см. Metabola (2)] состоит только из прементума и пары щупиков; глоссы и параглоссы утрачены; лабиальные щупики направлены дистально и сближены (так что их иногда принимают за глоссы), 2-члениковые [вместо исходных трехчлениковых – см. Hexapoda (3c)]; лишь у блох число члеников лабиального щупика вторично увеличено [см. ниже, Aphaniptera (1b)]. Возможно, отсутствие глосс и параглосс является синапоморфией Enteracantha и Diptera (см. «Систематическое положение» II).
(6) Особенности жилкования крыльев. Передние и задние крылья сходны по размеру, форме и жилкованию дистальной части, но имеют следующие различия в жилковании проксимальной части (рис. 10.9.1A, 10.11.1). На заднем крыле жилка Cu2 [исходно вогнутая и лежащая впереди клавального желоба или в клавальном желобе – см. Neoptera (1b)] и жилка Pcu [исходно выпуклая и лежащая сразу позади клавального желоба – см. там же] слиты на некотором протяжении, так что между местом их слияния и основанием крыла имеется ячейка, пересекаемая клавальным желобом. В результате этого проксимальный отрезок Pcu, результат слияния Pcu+Cu2 и дистальный отрезок Cu2 выглядят как единая продольная жилка, обрамляющая сзади клавальный желоб, а проксимальный отрезок Cu2 выглядит как отдельная короткая Y-образная жилка, дистально впадающая и в Cu1, и в Pcu. В отличие от заднего, на переднем крыле сохраняется исходное состояние: Cu2 и Pcu не слиты и разделены клавальным желобом. Нигмы [см. Metabola (5)] расположены одинаково на переднем и заднем крыле: две нигмы находятся между RS и M (одна проксимальнее первой поперечной жилки, вторая дистальнее), третья, наиболее проксимальная, нигма находится между Cu1 и Cu2. Такое расположение нигм позволяет гомологизировать жилки именно так, как указано здесь: наиболее проксимальная нигма (расположенная между Cu1 и Cu2) на переднем крыле находится в развилке между этими двумя ветвями Cu, а на заднем крыле она находится между Cu1 и сложной жилкой, дистальная часть которой соответствует Cu2; при этом на крыльях обеих пар эта проксимальная нигма отделена одним полем от того поля, где находятся две другие нигмы, гомология которых не вызывает сомнений.
По мнению некоторых авторов общий отрезок Cu2+Pcu на заднем крыле имеется также у Amphiesmenoptera и является апоморфией Panmecoptera (см. Mecopteriformia: «Классификация»); однако у Amphiesmenoptera место предполагаемого слияния жилок находится у самого основания крыла, плохо выражено или вообще не выражено (ср. рис. 10.16.1) и не отмечено нигмой, так что этот признак недостоверен.
Передние и задние крылья, будучи сходными по размеру и форме, в полете не сцеплены, т.е. насекомые бимоторные. Контактный аппарат крыльев [см. Metabola (4)] имеется, но не служит для механической сцепки передних и задних крыльев. Югальная доля переднего крыла слабо выступающая, несет одну или несколько длинных югальных щетинок, направленных назад-дистально; при расправленных крыльях эти щетинки налегают на передний край заднего крыла дистальнее плечевого выступа (у Merope/g1 эти щетинки слиты – см. ниже, Scorpiomuscae: «Классификация»); френулюм заднего крыла в виде одной или нескольких (часто двух) длинных щетинок, прижимающихся снизу к югуму.
Указанные здесь особенности строения крыльев достоверно относятся к Scorpiomuscae, Nannomecoptera и Metamecoptera; у всех Calyptroptera крылья вестигиальные и полностью лишены жилок, так что их исходное жилкование неизвестно [см. ниже, Calyptroptera (1)]. Крылья редуцированы также у некоторых других скорпионниц [см. ниже, Scorpiomuscae: «Классификация» и Metamecoptera (2)].
(7) Редукция яйцеклада. Яйцеклад самки не имеет по крайней мере придатков IX уромера [кинетапофизов и стилигеров – см. Amyocerata (7) и Eumetabola (1)]. Из тех частей, которые можно интерпретировать как первичные элементы яйцеклада, может сохранятся только пара вентральных склеритов – предположительно VIII кинетапофизы; эти склериты либо выступают позади стернита VIII уромера в виде пары выростов, либо соединены мембраной в непарный вырост, либо представляют собой пару склеритов на вентральной стенке VIII уромера, либо слиты в непарный склерит – вторичный стернит VIII уромера (рис. 10.8.2). Половое отверстие [исходно расположенное между стернитами VII и VIII уромеров – см. Amyocerata (7)] находится позади выростов VIII уромера, т.е. на границе VIII и IX уромеров. У Scorpiomuscae эти вентральные пластинки VIII уромера выглядят принадлежащими IX уромеру [см. ниже, Scorpiomuscae (1)]. У Boreus/fg2 они образуют вентральную створку яйцеклада, дорсальная створка которого образована IX и X уромерами (см. ниже, Raphioptera: «Классификация»).
(8) Гениталии. Гениталии самца включают генитальный тегмен [см. Metabola (6)], состоящий из пары двучлениковых парамеров, базальные членики которых (базипарамеры) соединены между собой дорсальным югумом (между кишечником и гонодуктом) и вентральным югумом (вентральнее гонодукта); весь тегмен соединен с тергитом и стернитом IX уромера так, что не способен в него втягиваться (в отличие от Hymenoptera и Eleuterata – рис. 10.1.4).
У скорпионниц (Scorpiomuscae, Metamecoptera, Nannomecoptera и Raphioptera) и дорсальный, и вентральный югумы вполне развиты [у Raphioptera смещены вперед – см. ниже, Calyptroptera (4)]; тегмен не способен втягиваться внутрь IX уромера, но при этом сохраняется хорошо видимый снаружи шов, отделяющий его от тергита и стернита IX уромера (рис. 10.9.1B, 10.9.2A, 10.10.2A, 10.11.1, 10.12.2D). У блох (Aphaniptera) вентральный югум разомкнут и превращен в пару аподем (манубриев); тегмен слит с тергитом IX уромера так, что шов между ними снаружи не виден; стернит IX уромера соединен с тегменом подвижно благодаря паре мыщелков (рис. 10.13.1D) [см. ниже, Aphaniptera (6a)].
К базипарамерам подвижно причленена пара телопарамеров; у Scorpiomuscae, Metamecoptera и Nannomecoptera телопарамеры двигаются навстречу друг другу, у Raphioptera и Aphaniptera – в дорсально-вперед [см. ниже, Calyptroptera (4)].
(9) Pupa dectica. Куколка исходно с подвижными мандибулами – «pupa dectica» [см. Metabola (1Bb)]. У куколки рострум [см. (2)] может быть короче, чем у имаго, а мандибулы длиннее имагинальных (рис. 10.10.2E). Развитые куколочные мандибулы имеются не только у представителей с развитыми имагинальными мандибулами (Scorpiomuscae, Metamecoptera, Raphioptera), но и у Nannomecoptera, у которых имагинальные мандибулы вестигиальные и неподвижны. Исключение составляют блохи (Aphaniptera), у которых мандибулы полностью утрачены и на стадии имаго, и на стадии куколки, так что куколки относятся к типу «pupa adectica» [см. ниже, Aphaniptera (8)].
Геологический возраст. Пермь – ныне (рис. 2.2) (Rasnitsyn & Quicke 2002). Начиная с нижней перми известны ископаемые насекомые с характерным для скорпионниц жилкованием и Y-образным проксимальным отрезком Cu2 на заднем крыле [см. Enteracantha (6)].
Систематическое положение Enteracantha. Помимо принятого здесь отнесения Enteracantha непосредственно к Mecopteriformia, предлагаются следующие классификации.
I. Объединение Enteracantha, Diptera и Amphiesmenoptera в таксон Panmecoptera (см. Mecopteriformia: «Классификации»).
II. Объединение Enteracantha и Diptera в таксон Antliophorida Weaver 1984. Близким по смыслу является объединение скорпионниц (Mecaptera) и Diptera в таксон Antliophora Hennig 1969, куда, в отличие от Antliophorida, не включены блохи (Aphaniptera). В качестве аутапоморфий Antliophorida и Antliophora называют следующие признаки (Hennig 1969, Mickoleit 1971).
(1) У самцов имеется семенной насос, накачивающий сперму из полового аппарата самца в половые пути самки. Однако среди скорпионниц семенной насос имеется только у Scorpiomuscae и его поршнем служит склерит, непосредственно сочлененными с внешним покровом генитального тегмена [см. ниже, Scorpiomuscae (3)]; в отличие от этого, у двукрылых камера и поршень насоса находятся в глубине генитального тегмена, на проксимальном конце эндофаллуса [см. Diptera (5b)]. Вероятно, семенные насосы Scorpiomuscae и Diptera возникли независимо, так же как независимо возникли разнообразные по строению семенные насосы других насекомых (см. Указатель признаков [3.58]).
(2) У имаго 3-й членик максиллярного щупика [5-членикового – см. Hexapoda (2c)] утолщен и несет комплекс сенсилл, расположенных в особом углублении. Этот признак выражен у Entracantha-Nannomecoptera (но не у других Enteracantha) и некоторых Diptera-Nemocera. Некоторые авторы считают его не аутапоморфией Antliophorida, а аутапоморфией филогенетической ветви, объединяющей только Nannomecoptera и Diptera (см. ниже, Nannomecoptera: «Систематическое положение»).
(3) У имаго глоссы и параглоссы утрачены, а лабиальные щупики сближены, направлены дистально и утратили исходное 3-члениковое строение [см. Enteracantha (5) и Diptera (1a)]. Неуникальный признак: утрата глосс и/или параглосс и сокращение числа члеников лабиальных щупиков многократно происходила у разных насекомых.
III. Объединение Mecaptera и Amphiesmenoptera в таксон Lepidopteroidea Escherich 1914. В качестве аутапоморфии Lepidopteroidea указывают то, что на заднем крыле Cu2 и Pcu слиты [см. Enteracantha (6)]; некоторые авторы считают этот признак апоморфией Panmecoptera или Mecopteria (см. Mecopteriformia: «Классификации»).
Статус Enteracantha. Традиционно скорпионниц (Mecaptera – см. «Классификация» I) и блох (Aphaniptera) рассматривают как два отряда, не объединяемые в таксон Enteracantha. Предлагались классификации, исключающие таксон Enteracantha: объединение Mecaptera и Diptera в таксон Antliophora (см. «Систематическое положение» II); объединение Mecaptera и Amphiesmenoptera в таксон Lepidopteroidea (см. «Систематическое положение» III); объединение Aphaniptera с Diptera в таксон Dipteroidea (см. ниже, Aphaniptera: «Систематическое положение» II).
Широко распространено представление о том, что отряд Mecaptera (= Mecoptera) является предковым не только для Aphaniptera, но и для Diptera и/или для Amphiesmenoptera. В связи с этим название таксона, объединяющего отряды Mecaptera, Diptera, Trichoptera и Lepidoptera, принято образовывать от того или иного названия скорпионниц – Panorpoidea (sensu Handlirsch 1903), Mecopteroidea auct., Panmecoptera Crampton 1924, Mecopteria Hennig 1953, Mecopteriformia Ax 1999 и др. В этом случае предполагается парафилия не только таксона Mecaptera, но и таксона Enteracantha в целом. Однако таксон Mecaptera не может быть предковым для бабочек и ручейников, потому что у скорпионниц ноги личинок исходно претерпели необратимую редукцию [см. Enteracantha (4)], тогда как у бабочек и ручейников личиночные ноги исходно сохраняют примитивное строение [ср.: Amphiesmenoptera (13)]. Представление о происхождении двукрылых от скорпионниц также представляется неверным, потому что у двукрылых, как и у бабочек и ручейников, отсутствует аканты провентрикулюса, при том что эти аканты консервативно сохраняются у скорпионниц и блох [см. Enteracantha (1)].
Классификация Enteracantha
I. Деление Enteracantha на Mecaptera и Aphaniptera. Таксон Mecaptera Packard 1886a, 1886b (synn.circ.: Mecoptera Hyatt & Arms 1890 = Eumecoptera Tillyard 1919b = Mecopterida Pearce 1936 = Mecopterodea Boudreaux 1979b) объединяет всех Enteracantha кроме блох (Aphaniptera); он представляет собой плезиоморфон и характеризуется лишь отсутствием приспособлений к кровососанию и прочих апоморфий, свойственных блохам. В отличие от блох, большинство Mecaptera крылатые; но имеются короткокрылые и бескрылые представители [см. Enteracantha (6)]. В отличие от блох, многие Mecaptera имеют хорошо развитый рострум; но у некоторых представителей рострум укорочен или совсем не выражен [см. Enteracantha (2)]. В отличие от блох, у большинства Mecaptera ротовой аппарат грызущий, с развитыми мандибулами; но у наннохорист сосущий [см. Enteracantha (2)]. В отличие от блох, куколки которых неподвижны и не имеют мандибул, куколки всех изученных в этом отношении Mecaptera имеют функционирующие мандибулы (pupa dectica).
Принято считать, что отряд Mecaptera (= Mecoptera) является парафилетическим, предковым как для блох, так и для двукрылых (Diptera), и/или для бабочек и ручейников (Amphiesmenoptera); последнее утверждение ошибочно (см. выше, «Статус»).
II. Деление Enteracantha на Calyptroptera и всех остальных. Некоторые авторы объединяют Scorpiomuscae, Metamecoptera и Nannomecoptera в таксон «Mecoptera» sensu Hinton 1958 (non Hyatt & Arms 1891). Эти таксоны имеют некоторые общие признаки неясного филогенетического значения (Hinton 1958), в том числе следующие. У имаго простые глазки сближены (тогда как у Raphioptera они широко расставлены, а у Aphaniptera отсутствуют). У имаго самца телопарамеры [см. Enteracantha (8)] сгибаются навстречу друг другу (тогда как у Calyptroptera они сгибаются дорсально-вперед). Оварии политрофические (тогда как у Calyptroptera паноистические).
III. Выделение таксона Promecoptera. Таксон Promecoptera Crampton 1916a либо включает только Merope/fg, либо также Notiothauma/fg (см. ниже, Scorpiomuscae: «Классификация»). Часто этот таксон фигурирует под неверным названием «Protomecoptera» sensu Crampton 1930 (что не соответствует исходной циркумскрипции таксона Protomecoptera Tillyard 1917c, установленного для единственного вымершего вида Archipanorpa magnifica Tillyard 1917). Promecoptera противопоставляют всем прочим Mecaptera, объединяемым под ошибочным названием «Eumecoptera» auct. (non Eumecoptera Tillyard 1919b; nec Eumecoptera Crampton 1930). Эта классификация основана лишь на том, что у Promecoptera крылья широкие, с густым жилкованием и с многочисленными жилками в костальном поле; при этом считают, что у других скорпионниц произошло уменьшение числа жилок и сужение крыла. Данное представление об эволюции скорпионниц основано не на анализе уникальных апоморфий, а на необоснованном «законе» олигомеризации органов (см. I.4.2.4).
IV. Выделение таксона Pistillifera. В этой классификации таксон Nannomecoptera противопоставляется всем прочим скорпионницам.
Таксон Pistillifera Willmann 1987 включает всех Mecaptera кроме Nannomecoptera (см. ниже) и характеризуется тем, что у самца исходно имеется семенной насос, прикрепленный к генитальному югуму [см. ниже, Scorpiomuscae (3)] (Willmann 1987). Однако у Raphioptera генитальный югум удален от пениса [см. ниже, Calyptroptera (4)], так что он не может служить местом прикрепления насоса.
Таксон Opisthogonophora Willmann 1987, объединяет всех Pistillifera кроме Metamecoptera и характеризуется тем, что половое отверстие самки открывается позади IX стернита брюшка (тогда как у Nannomecoptera оно открывается позади VIII стернита). Однако у Raphioptera и Metamecoptera стернит IX сегмента брюшка отсутствует, так что этот признак не выражен (рис. 10.8.2).
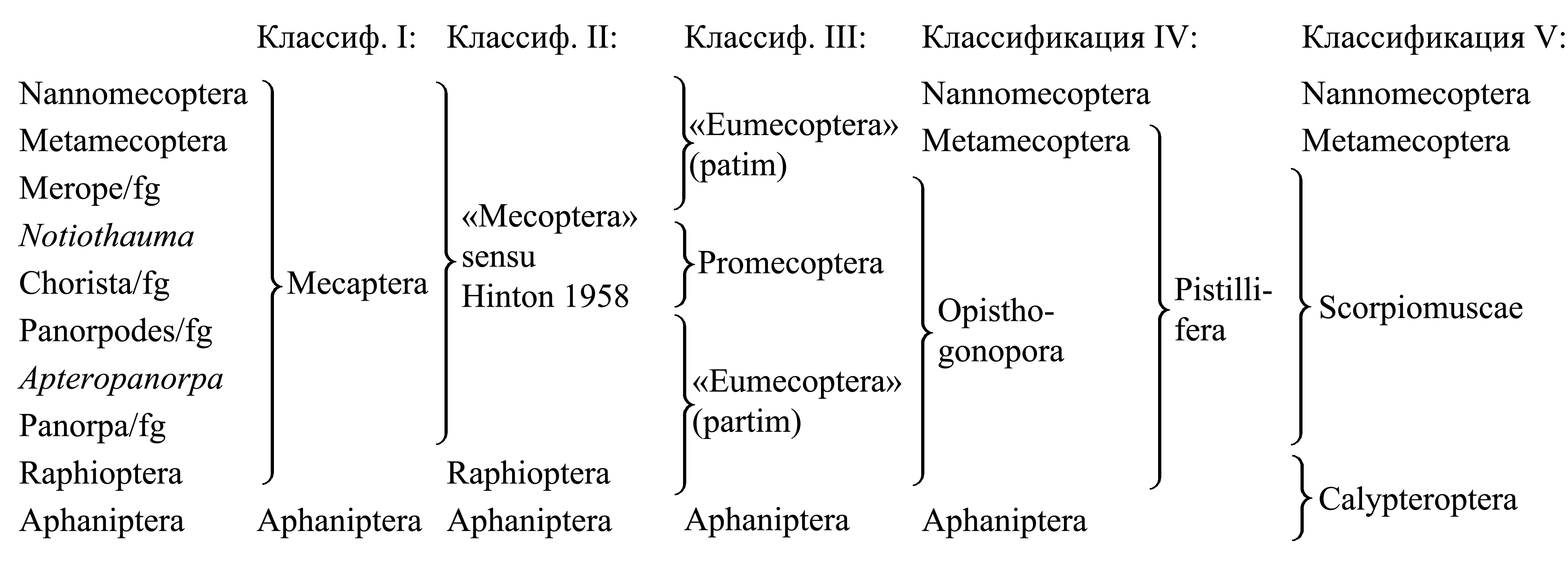
V. Здесь мы рассматриваем в составе Enteracantha четыре непосредственно подчиненных таксона – Scorpiomuscae, Metamecoptera, Nannomecoptera и Calyptroptera, который делится на Raphioptera и Aphaniptera.

Enteracantha, или Panorpa/fg1
![]() 1. Scorpiomuscae,
или Panorpa/fg2
1. Scorpiomuscae,
или Panorpa/fg2
![]() 2. Metamecoptera,
или Bittacus/fg1
2. Metamecoptera,
или Bittacus/fg1
![]() 3. Nannomecoptera,
или Nannochorista/fg1
3. Nannomecoptera,
или Nannochorista/fg1
![]() 4. Calyptroptera,
или Pulex/fg1
4. Calyptroptera,
или Pulex/fg1
![]() 4.1. Raphioptera,
или Boreus/fg1
4.1. Raphioptera,
или Boreus/fg1
![]() 4.2. Aphaniptera,
или Pulex/fg2
4.2. Aphaniptera,
или Pulex/fg2