КЛАДОЭНДЕЗИС НАСЕКОМЫХ

![]()
|
КЛАДОЭНДЕЗИС НАСЕКОМЫХ |
|||||||
|
|
|
fg | |||||
IX-1.2.1. Panpsocoptera Crampton 1938 –
(Somobialia![]() Eucaryota
Eucaryota![]() Metazoa
Metazoa![]() Monostomata
Monostomata![]() Haemataria
Haemataria![]() Peritoneata
Peritoneata![]() Metameria
Metameria![]()
Gnathopoda![]() Euarthropoda
Euarthropoda![]() Mandibulata
Mandibulata![]() Atelocerata
Atelocerata![]() Hexapoda
Hexapoda![]() Amyocerata
Amyocerata![]()
Pterygota![]() Metapterygota
Metapterygota![]() Neoptera
Neoptera![]() Eumetabola
Eumetabola![]() Parametabola
Parametabola![]() Acercaria
Acercaria![]() Panpsocoptera)
Panpsocoptera)
(рис. 9.2.1–9.3.5)
(рис. 9.2.1,
9.2.2,
9.2.3, 9.2.4,
9.2.5,
9.2.6,
9.3.1,
9.3.2,
9.3.3,
9.3.4,
9.3.5)
|
Названия, однозначно соответствующие по циркумскрипции: — Panpsocoptera = Psocopteria Crampton 1938; — Psocopteroida Müller 1963. По циркумскрипции также соответствует таксонам под ошибочными типифицированными названиями, образованными от родового названия Psocus – Psociformia, Psocoidea, Psocodea, Psocodida и др. Иерархическое название: Pediculus/f11=g1 [f:1777; g:L.1758(gen.233)]. |
|
Ранг. Обычно надотряд.
Название. Наиболее ранним нетипифицированным названием таксона, объединяющего Copeognatha и Parasita, является Eupsocoptera Lameere 1936 (исходно во французском написании, как Eupsocoptères). Однако исходно в состав Eupsocoptera, помимо современных таксонов был включен вымерший таксон Tetrastigmoptera, который ошибочно относили к Copeognatha (см. Condylognatha: «Систематическое положение»). Таким образом, название Eupsocoptera не соответствующим по циркумскрипции таксону, называемому здесь Panpsocoptera.
Аутапоморфии Panpsocoptera, выраженные у Copeognatha и Parasita. Данные апоморфии служат доказательством того, что Copeognatha и Parasita, традиционно считающиеся разными отрядами, образуют голофилетический таксон Panpsocoptera.
(1) Модификации глотки и гипофаринкса (рис. 9.2.3, 9.2.4C). Кутикулярная выстилка вентральной, или задней, стенки глотки (которая дистальнее переходит в дорсальную стенку гипофаринкса) образует ситофорный склерит, называемый также глоточным, или фарингеальным склеритом [«ситофором» называют обычное для всех насекомых медиальное углубление в основании дорсальной стенки гипофаринкса, переходящее в вентральную выстилку глотки (Snodgrass 1944)]. Ситофорный склерит представляет собой цельный крупный непарный склерит сложной формы, медиальная часть которого имеет вогнутую чашевидную форму. На противоположной стенке глотки (дорсальной, или передней) имеется выступающий цибариальный склерит, который служит местом прикрепления мощных цибариальных мышц, идущих от увеличенного клипеуса [см. Acercaria (4)].
На вентральной (задней) стороне гипофаринкса (т.е. на стороне, обращенной к нижней губе) имеется пара гипофарингеальных пластинок. Каждая гипофарингеальная пластинка представляет собой хорошо очерченный участок покрова овальной формы, тянущийся по вентральной стороне гипофаринкса от основания до вершины; от ее проксимального конца, который находится у основания гипофаринкса (т.е. в углублении между гипофаринксом и нижней губой), отходит мышца, идущая к заднему краю головной капсулы.
Ситофорный склерит и пара гипофарингеальных пластинок соединены Y-образным гипофарингеальный лигаментом; это кутикулярный тяж, лежащий под покровами; он начинается в виде непарного тяжа от глоточного склерита и тянется в дистальном направлении под дорсальной (передней) стенкой гипофаринкса; там он разделяется на пару ветвей; каждая ветвь, обогнув вершину гипофаринкса, переходит на его вентральную (заднюю) сторону и там идет по гипофарингеальной пластинке в проксимальном направлении. У некоторых Parapsocida гипофарингеальный лигамент не Y-образный, а на всем протяжении парный. У некоторых сеноедов и пухоедов гипофарингеальные пластинки являются железами, а лигаменты представляют собой каналы с просветом внутри. У других сеноедов и пухоедов гипофарингеальные пластинки склеротизованы, не железистые, а лигаменты не имеют просвета. Некоторые авторы предполагали какую-то связь между гипофарингеальными пластинками и слюнными железами; однако у сеноедов и пухоедов, имеющих гипофарингеальные пластинки, слюнной проток [непарный, открывающийся в углублении между гипофаринксом и нижней губой – см. Hexapoda (3)] не имеет связи с гипофарингеальными пластинками.
Наличие гипофарингеальных пластинок и их лигаментов – уникальная апоморфия; они имеются у всех сеноедов (Parapsoicida и Eupsocida) и у всех пухоедов (Amblycera и Ischnocera); у Rhyncophthirina с их вестигиальным гипофаринксом гипофарингеальные пластинки утрачены; у сосущих вшей гипофаринкс крайне видоизменен [см. ниже, Siphunculata (1e)].
Помимо модификации глотки, ротовой аппарат имеет особенности, свойственные Acercaria: максиллы лишены кардо, с подвижной малой и втягивающейся лацинией [см. Acercaria (1) и рис. 9.2.4A–B, 9.3.1B]. При этом у сеноедов и пухоедов (Parapsocida, Eupsocida и Mallophaga) ротовой аппарат сохраняет грызущую специализацию: мандибулы вполне развиты, а лациниальные малы приспособлены для перемещения в рот частиц пищи, измельченной мандибулами. Втягивающиеся лацинии максилл не образуют сосательного аппарата (в отличие от Condylognatha); они либо обеспечивают движение мал (у Eupsocida и, вероятно, также у Parapsocida), либо вестигиальные и не функционируют (у Mallophaga) [см. ниже, Parapsocida (2), Eupsocida (2), Parasita (5)]. Таким образом, особое строение глотки и гипофаринкса не связано с изменением специализации ротового аппарата. Ротовые аппараты Siphunculata и Rhyncophthirina изменили специализацию с грызущей на сосательную, но одновременно с этим утратили особенности глотки и гипофаринкса, исходные для Panpsocoptera.
(2) Гениталии самца. У самца копулятивный аппарат, или пенис [находящийся между IX стернитом брюшка и парапроктами – см. Amyocerata (6)] отчасти мембранозный и обрамлен склеритом (называемым базальной пластинкой, или рамкой пениса), так что непарный семенной канал проходит сквозь эту склеротизованную рамку; часть рамки вентральнее семенного канала вдается в карман, так что скрыта IX стернитом брюшка и служит вентральной аподемой пениса; от нее к IX стерниту идут мышцы, способные выдвигать пенис наружу (рис. 9.2.1B). Дистально (сзади) рамка пениса обычно несет пару отростков; иногда их называют парамерами, но, в отличие от парамеров Condylognatha они не имеют собственных мышц и сочленений. Придатки, сопоставимые с парамерами Condylognatha, у Panpsocoptera отсутствуют.
(3) Редукция яйцеклада. У самки яйцеклад [см. Amyocerata (7) и Eumetabola (1)] уменьшен и не выполняет функцию проведения яиц (рис. 9.2.1A, 9.2.6). У Parasita яйцеклад подвергся дальнейшей редукции: его кинетапофизы утрачены, так что имеются лишь IX стилигеры в виде пары небольших широко расставленных неподвижных отростков; у Amblycera и части Siphunculata они также утрачены.
(4) Модификация десятого сегмента брюшка. Тергит X уромера [последнего – см. Hexapoda (8) и Amyocerata (8)] уменьшен так, что весь расположен дорсальнее парапроктов и между ними, не заходя на бока тела впереди от парапроктов, так что представляет собой лишь конический выступ (называемый здесь «эпипрокт»), по размеру сопоставимый с парапроктами (рис. 9.2.1A). У крылатых имаго (имеющихся только среди сеноедов – Parapsocida и Eupsocida) каждый парапрокт несет полусферическую выпуклость с трихоботриями (рис. 9.2.1B). У некоторых Parasita IX и X сегменты брюшка слиты.
Аутапоморфии Panpsocoptera, выраженные только у Copeognatha. Данные апоморфии выражены только у крылатых форм, имеющихся только среди сеноедов (Copeognatha) – как Parapsocida, так и Eupsocida; у всех Parasita крылья утрачены и указанные ниже особенности отсутствуют.
(5) Крыловая зацепка. Зацепка, обеспечивающая сцепление заднего крыла с передним в полете [см. Parametabola (2)], имеет особое строение (рис. 9.2.1C, 9.2.5D). На переднем крыле по самому дну клавального желоба [вогнутого дорсально и выпуклого вентрально – см. Neoptera (1b)] проходит жилка, обычно обозначаемая как Cu2; эта жилка эластична и армирована экзотенидиями [см. Pterygota (4Ae)]; около места впадения Cu2 в задний край крыла (т.е. около заднего нодуса) экзотенидии на ее выпуклой вентральной стороне вытянуты в выступающие зубцы, направленные в сторону заднего края крыла. Этот пучок зубцов у вершины Cu2 называют нодулюс (nodulus); у Parapsocida нодулюс состоит из нескольких тонких зубцов (рис. 9.2.5D), у Eupsocida он представляет собой один крупный изогнутый зубец (рис. 9.2.1С) [см. ниже, Parapsocida (1) и Eupsocida (1)]. Участок заднего края крыла позади нодулюса завернут вентрально и вперед, так что направлен навстречу зубцам нодулюса. На заднем крыле костальный край завернут дорсально; он зацепляется за подвернутый задний край переднего крыла и удерживается зубцами нодулюса (рис. 9.2.1С). В отличие от Thysanoptera и Plantisuga, у Panpsocoptera нет крючковидных сцепочных щетинок на заднем крыле. Сходная зацепка с пучком зубцов на переднем крыле независимо возникла у Heteropteroidea; но там зубцы сидят не на жилке, идущей по клавальному желобу, а на клавусе, и костальный край заднего крыла изогнут S-образно [ср.: Heteropteroidea (1)].
На переднем крыле, помимо зацепки около заднего края, функционирующей при полете, имеется вторая зацепка, расположенная около переднего края крыла и функционирующая при сложенных крыльях – это стигмапофиз (stigmapophysis), называемый также нодус (nodus). Стигмапофиз находится на вентральной стороне крыла на жилке RA и направлен назад; у Parapsocida стигмапофиз состоит из нескольких тонких зубцов, т.е имеет такое же строение, как и нодулюс на том же крыле [см. ниже, Parapsocida (1) и рис. 9.2.5C]; у Eupsocida стигмапофиз представляет собой цельный выступ, расположенный около проксимального края птеростигмы [см. ниже, Eupsocida (1) и рис. 9.2.1A)]. При сложенных крыльях костальный край заднего крыла упирается в стигмапофиз переднего крыла.
У некоторых сеноедов с упрощенным строением крыльев зацепка и/или стигмапофиз утрачены (рис. 9.2.6). У некоторых сеноедов (некоторые Parapsocida и самки некоторых Eupsocida) и у всех Parasita крылья полностью утрачены. Можно предположить, что особенности строения крыльев, общие для крылатых Parapsocida и крылатых Eupsocida, являются исходными для Panpsocoptera. В этом случае наличие компактной зацепки на переднем крыле является аутапоморфией Panpsocoptera.
Помимо строения зацепки и стигмапофиза, крылья сеноедов имеют некоторые другие особенности, предположительно исходно присущие Panpsocoptera (рис. 9.2.1, 9.2.5). Жилка Sc не выражена. Жилка RS отходит от RA и обычно разделяется на две ветви, впадающие в передний край крыла. Жилки RS и M соединены либо посредством постоянной поперечной жилки, либо слившись в одной точке, либо слившись на некотором протяжении (рис. 9.2.1A) (то же у Zoraptera). Жилки M и Cu1 имеют общее основание и разделяются проксимальнее места соединения M с RS. Жилка M обычно разделяется на 3 или 4 ветви, впадающие в край крыла позади его вершины. «Areola postica» [см. Parametabola (3)] бывает выражена: в этом случае Cu1 бывает двуветвистая, а ее передняя ветвь Cu1a выгнута вперед (рис. 9.2.1A); выгнутая Cu1a может соединяться с M посредством постоянной поперечной жилки, либо сливаясь с ней на некотором протяжении. Иногда «areola postica» не выражена: Cu1a бывает прямой (рис. 9.2.5A), либо отсутствует (рис. 9.2.6). На клавусе обычно имеется единственная жилка Pcu (рис. 9.2.1A), либо 2 жилки – Pcu и V. На переднем крыле Pcu впадает либо в задний край клавуса, либо в вершину жилки Cu2 в области нодулюса (см. выше); на заднем крыле Pcu впадает в задний край клавуса. У многих видов часть названных жилок отсутствует. На заднем крыле жилок меньше, чем на переднем.
(6) Орган Пеармана. На задних ногах у имаго обоих полов (но не у личинок) бывает развит стридуляционный орган Пеармана (Pearman 1928): на внутренней стороне каждого из задних тазиков находятся расположенные рядом терка (бугорок, покрытый морщинками или зубчиками) и зеркальце (небольшая мембранозная площадка). Орган Пеармана развит у многих крылатых видов сеноедов – как Parapsocida, так и Eupsocida; у некоторых видов терка и/или зеркальце отсутствуют. У бескрылых сеноедов орган Пеармана утрачен; его нет также у Parasita.
(7) Прядильные железы. Лабиальные железы [открывающиеся общим непарным отверстием между гипофаринксом и нижней губой – см. Hexapoda (3)] частично специализированы для выделения шелка: в общее отверстие или в общий проток открываются протоки от двух пар лабиальных желез; из них более дорсальная пара выделяет шелк, тогда как более вентральная пара сохраняет функцию слюнных желез (Weber 1938). Выделение шелка свойственно многим видам сеноедов (Parapsocida и Eupsocida), но некоторые виды сеноедов этой способность не обладают. У Parasita выделение шелка не наблюдается, но имеются две пары лабиальных желез, существенно отличающихся друг от друга по строению и функциям (Благовещенский 1959). Самки сеноедов используют шелк для оплетения кладки яиц; иногда шелком оплетаются также гнезда, в которых живут колонии личинок и имаго сеноедов. Помимо Panpsocoptera, специализация лабиальных желез в качестве шелкоотделительных независимо возникла у Mecopteriformia.
Геологический возраст. Юра – ныне. Наиболее древние достоверные представители Parapsocida и Eupsocida известны из юры Казахстана (Rasnitsyn & Quicke 2002). Более точно определить возраст Panpsocoptera трудно, поскольку для большинства ископаемых сеноедоподобных насекомых известно только строение крыльев и неясно, относятся ли они к сеноедам или к вымершему таксону Tetrastigmoptera. Иногда указывают, что сеноеды (Copeognatha) известны начиная с перми; в этом случае в состав отряда Copeognatha включают Tetrastigmoptera (см. Condylognatha: «Систематическое положение»).
Классификации Panpsocoptera
I. Деление Panpsocoptera на Haplognatha и Siphunculata. Таксон Haplognatha Börner 1904a объединяет сеноедов (Copeognatha) и пухоедов (Mallophaga); он характеризуется уникальными особенностями строения глотки и гипофаринкса [см. Panpsocoptera (1)], тогда как у вшей (Siphunculata), а также у слоновьей вши (Rhyncophthirina), эти особенности отсутствуют. Это объясняется тем, что ротовые аппараты Siphunculata и Rhyncophthirina настолько видоизменены, что не сопоставимы ни с ротовым аппаратом Haplognatha, ни с ротовыми аппаратами других насекомых [см. ниже, Siphunculata (1) и Rhyncophthirina (1)]. Судя по всему, таксон Haplognatha является парафилетическим, предковым и для Siphunculata, и для Rhyncophthirina.
II. Деление Panpsocoptera на Copeognatha и Parasita. Таксон Parasita объединяет Mallophaga, Siphunculata и Rhyncophthirina, характеризуется аутапоморфиями и является голофилетическим (см. ниже, Parasita).
Плезиоморфон Copeognatha Enderlein 1903 – сеноеды {synn.circ: Psocoptera Shipley 1904 = Clinoptera Crampton 1916a = Corrodentida Pearce 1936; по циркумскрипции также соответствует: «Corrodentia» sensu Handlirsch 1903 (non Burmeister 1839); типифицированные названия: Psocus/fg [f:1810; g:1794] (употребляются названия Psocus, Psoquillae, Psocides, Psocina, Psocidae, Psocidi, Psocoidea, Psocida и др.)}. Этот таксон широко признан в ранге отряда и характеризуется следующими плезиоморфиями. Ротовой аппарат грызущий, с вполне развитыми грызущими мандибулами. Втягивающиеся лацинии максилл [см. Acercaria (1)] всегда хорошо развиты, функционирующие, никогда не подвергаются редукции или уменьшению (в отличие от всех Parasita). Максиллярный щупик всегда 4-члениковый [см. Acercaria (2)]. У гипофаринкса ситофорный склерит, гипофарингеальные пластинки и соединяющие их лигаменты всегда развиты [то же у Mallophaga – см. Panpsocoptera (1)]. Имеется нижняя губа с короткими 2-члениковыми или 1-члениковыми щупиками [см. Acercaria (3)]. Гула всегда отсутствует (в отличие от всех Parasita), так что шейная мембрана начинается сразу позади задних тенториальных ямок и основания нижней губы независимо от того, является ли голова гипогнатной (рис. 9.2.1A) или прогнатной (рис. 9.2.6). Передние тенториальные ямки [см. Amyocerata (3)] занимают передне-латеральное положение по бокам клипеуса и видны сверху (как у большинства насекомых, но в отличие от всех Parasita). Клипеус всегда обращен вперед и имеет характерную сильно выпуклую полушаровидную форму, связанную с наличием крупной непарной конической клипео-глоточной мышцы [см. Panpsocoptera (1)] (рис. 9.2.3). Антенны длинные, нитевидные: флагеллум антенны [см. Amyocerata (1)] с большим фиксированным числом члеников; флагелломеры удлиненные, тонкие и параллельносторонние; в отличие от флагелломеров, скапус и педицеллус не удлинены (рис. 9.2.1A); у личинки число члеников флагеллума меньше, чем у имаго, и увеличивается с возрастом (в отличие от всех Parasita). Крылья бывают вполне развиты, либо вестигиальные, либо утрачены (в отличие от Parasita, у которых крылья всегда полностью утрачены). В отличие от Parasita, сеноеды либо ведут образ жизни, не связанный с позвоночными животными, либо обитают в гнездах или норах птиц и зверей, но при этом не питаются ни кровью, ни покровами живых животных.
Таксон Copeognatha представляет собой плезиоморфон, предковый для голофилетического таксона Parasita. В составе Copeognatha принято выделять подчиненные таксоны «Troctomorpha», «Trogiomorpha» и Eupsocida (= Psocomorpha), либо «Troctomorpha» и «Trogiomorpha» объединяют в таксон Parapsocida (см. ниже, Parapsocida: «Классификация»). Подавляющее большинство сеноедов относится к таксону Eupsocida, который характеризуется аутапоморфиями и является голофилетическим [см. ниже, Eupsocida (1)–(2)]. Таксон Parapsocida представляет собой плезиоморфон, предковый и для Eupsocida, и для Parasita. Об этом, помимо прочих признаков, свидетельствует мускулатура максилл. У Parapsocida максилла имеет оба механизма сгибания малы – и лациниальный механизм, и мышцу, идущую к мале из базистипеса [см. Acercaria (1b)]. У Eupsocida сохраняется только лациниальный механизм, а у Parasita – только мышца, идущая из базистипеса. Поскольку эти насекомые имеют грызущие мандибулы и при этом их максиллы не имеют подвижных кардо, они способны питаться только при наличии какого-нибудь из этих механизмов сгибания малы. Если исходить из того, что утрата каждого из двух этих механизмов является необратимым эволюционным процессом, а также допустить, что она произошла однократно, то приведенная ниже филогенетическая схема представляется наиболее правдоподобной.
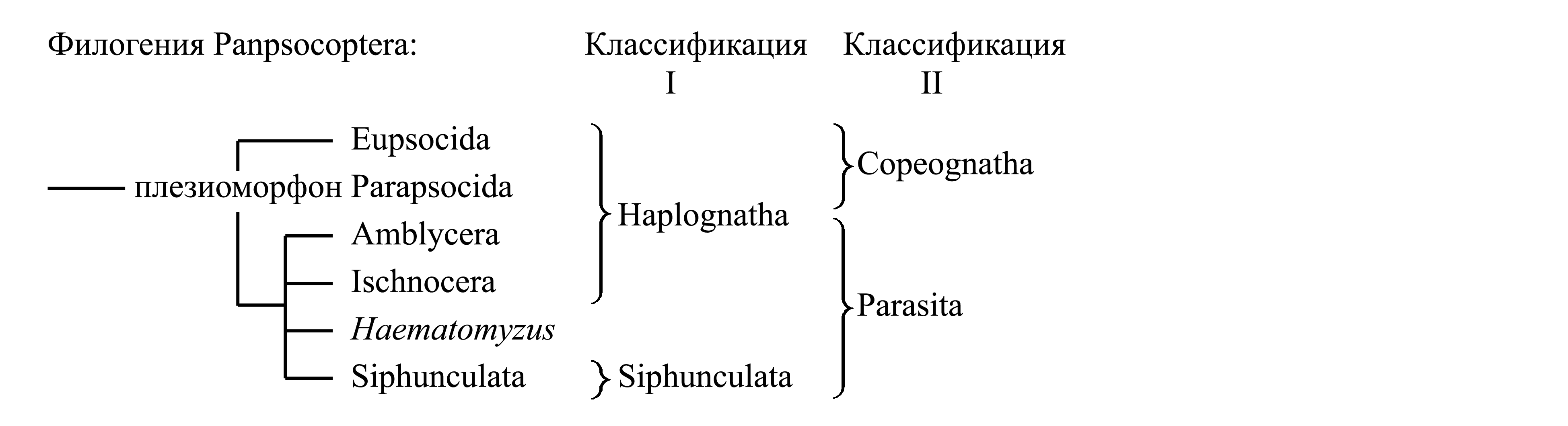
III. Деление Panpsocoptera на Parapsocida, Eupsocida и Parasita. Здесь мы принимаем деление Panpsocoptera на 3 непосредственно подчиненных таксона – плезиоморфон Parapsocida и два голофилетических таксона – Eupsocida (стр. 0000) и Parasita (стр. 0000).
Panpsocoptera, или Pediculus/f11=g1
![]() 1. Плезиоморфон Parapsocida, или
Atropos/f1=Trogium/g1
1. Плезиоморфон Parapsocida, или
Atropos/f1=Trogium/g1
![]() 2. Eupsocida, или Psocus/fg1
2. Eupsocida, или Psocus/fg1
![]() 3. Parasita, или Pediculus/f12=g2
3. Parasita, или Pediculus/f12=g2
![]() 3.1. Mallophaga, или
Philopterus/f1=Ricinus/g1
3.1. Mallophaga, или
Philopterus/f1=Ricinus/g1
![]() 3.1.1. Amblycera, или Ricinus/f1=g2
3.1.1. Amblycera, или Ricinus/f1=g2
![]() 3.1.2. Ischnocera, или
Philopterus/f2=g1
3.1.2. Ischnocera, или
Philopterus/f2=g1
![]() 3.2. Siphunculata, или
Pediculus/f13=g3
3.2. Siphunculata, или
Pediculus/f13=g3
![]() 3.3. Rhyncophthirina, или
Haematomyzus/fg(1)
3.3. Rhyncophthirina, или
Haematomyzus/fg(1)